




Факторы роста
Классификация и характеристики
-
Фактор роста тромбоцитов (PDGF)
PDGF состоит из двух подединиц, известных как цепочка А (18 кDa) и цепочка В (12-14 кDa), которые разделяют очень гомологичные пептидные последовательности, включая 8 консервированных остатков цистеина в идентичных положениях. PDGF существует в 3 изоформах: гомодимеры PDGF-AA и PDGF-BB и гетеродимер PDGF-AB. Эти изоформы распределяются по различным тканям и клеточным линиям, что предполагает разные биологические функции для каждой.
На поверхности клетки присутствуют два типа рецепторов PDGF: PDGFR-α и PDGFR-β. PDGFR-α может связывать как A, так и B подединицы PDGF, в то время как PDGFR-β связывает только B подединицу. PDGF-AA связывается с α-димерами рецепторов, PDGF-AB может образовать α- и β-димеры, а PDGF-BB может активировать α-, β- и β-димеры рецепторов. Эти димеры рецепторов представляют собой активные сигнальные формы PDGFR [1].
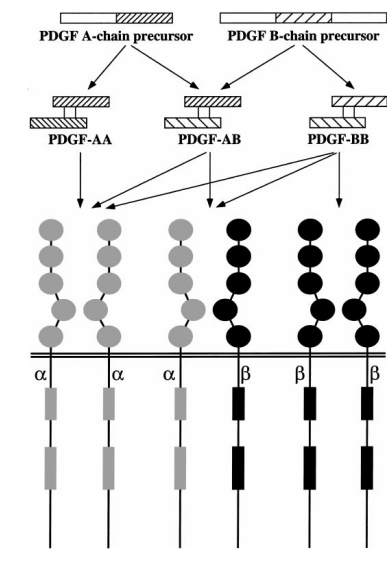
Рисунок 1: Обработка и функции изоформ фактора роста тромбоцитов (PDGF) [2]
-
Фактор роста сосудистой эндотелии (VEGF)
Фактор роста сосудистой эндотелии (VEGF), также известный как фактор сосудистой проницаемости (VPF), является высокоспецифическим митогеном для сосудистых эндотелиальных клеток. Семейство VEGF включает в себя несколько членов: VEGF-A, VEGF-B, VEGF-C, VEGF-D, VEGF-E и фактор роста плаценты (PlGF). Среди них, VEGF-A существует в четырех основных изоформах - VEGF121, VEGF165, VEGF189 и VEGF206, полученных через альтернативное сплейсинг мРНК.
VEGF оказывает свое биологическое воздействие, связываясь с тремя основными типами рецепторов тирозинкиназы: VEGFR-1 (Flt-1), VEGFR-2 (Flk-1/KDR) и VEGFR-3 (Flt-4). Каждая изоформа VEGF демонстрирует различные связывающие свойства к этим рецепторам, что способствует разнообразию сигнальных путей, опосредованных VEGF [3].
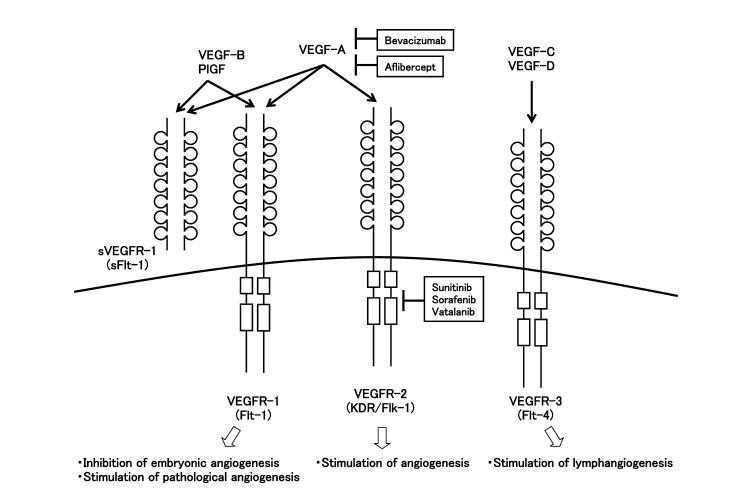
Рисунок 2: Члены семьи VEGF и их рецепторы
VEGF-A обычно называют просто VEGF и служит ключевым регулятором васкулогенеза развития, ангиогенеза и дифференциации эндотелиальных клеток-предшественников. VEGF-B играет роль в опухолях, где неоваскуларизация отсутствует. VEGF-C и VEGF-D участвуют в формировании как новых кровеносных сосудов, так и лимфатических сосудов в опухолевых тканях. VEGF-E также считается потенциальным ангиогенным фактором, в то время как PlGF (фактор роста плаценты) способствует неоваскуларизации и повышает сосудистую проницаемость.
-
Эпидермальный фактор роста (EGF)
Эпидермальный фактор роста (EGF) - небольшой полипептид, широко присутствующий у людей и других животных, состоящий из пептидной цепи с 50-60 аминокислотами. Эта цепочка содержит 6 остатков цистеина, которые образуют стабильные дисульфидные связи, в результате чего образуются 3 петельных домена, критически важных для его биологической активности. Даже при крайне низких концентрациях ЭГФ может сильно стимулировать рост клеток, ингибировать экспрессию генов, связанных со старением, и задерживать старение эпидермальных клеток.
EGF осуществляет свои действия, связываясь с внеклеточной областью (включающей домены I-IV) своего рецептора, EGFR (рецептор эпидермального фактора роста), приводя к димеризации рецептора тирозинкиназы [4]. Эта активация вызывает каскад биохимических событий, включая повышенный внутриклеточный уровень кальция, усиленный гликолиз и синтез белков, а также увеличенную экспрессию определенных генов, в том числе самого гена EGFR. В конечном счете, эти события способствуют синтезу ДНК и пролиферации клеток.
-
Факторы роста фибробластов (FGFs)
Факторы роста фибробластов (FGFs) - это семейство полипептидных факторов роста, присутствующих в различных тканях по всему организму. Они существуют в основном в двух тесно связанных формах: основной FGF (bFGF) и кислотный FGF (aFGF). Семейство FGF состоит из 22 лигандов, которые могут взаимодействовать с четырьмя различными рецепторами FGF (FGFR). Сигнальный путь FGF/FGFR регулирует фундаментальные клеточные процессы, такие как выживание клеток, пролиферация, миграция, дифференциация, развитие эмбриона, органогенез, восстановление / регенерация тканей и метаболизм.
Имя “ фактор роста фибробластов” отражает их мощное митогенное воздействие на фибробласты. Кроме того, FGF также стимулируют пролиферацию гладких мышечных клеток, хондроцитов, кератиноцитов и перицитов. Из-за их сильной аффинности к гепарину, FGF также известны как факторы роста, связывающие гепарин. Они синтезируются сосудистыми эндотелиальными клетками и хранятся в нижней мембране и внеклеточной матрице.
-
Инсулиноподобные факторы роста (IGF)
Семейство инсулиноподобных факторов роста (IGF) включает в себя 2 полипептида низкого молекулярного веса (IGF-1 и IGF-2), 2 типа специфических рецепторов (IGF-1R и IGF-2R, также называемых рецепторами типа I и типа II) и 6 связывающих IGF белков (IGFBPs). IGF-1R имеет структурное сходство с инсулиновым рецептором (IR), образуя гетеротетрамерный гликопротеин (α). ₂β₂) Состоит из 2 α-подединиц и 2 β-подединиц. Альфа-субединица содержит домен связывающий лиганды, в то время как β-субединица обладает внутренней активностью тирозинкиназы (но без активности тирозиназы).
Связывающие свойства инсулина и ИГФ к их соответствующим рецепторам варьируются:
- Для инсулинового рецептора (ИР): Инсулин > ИГФ-1 > ИГФ-2
- Для рецептора ИГФ-1 (ИГФ-1Р): ИГФ-1 > ИГФ-2 > Инсулин
- Для рецептора ИГФ-2 (ИГФ-2Р): ИГФ-2 > IGF-1, без перекрестной реакции на инсулин.
IGF-1 является одноцепным базовым белком, состоящим из 70 аминокислот, с молекулярной массой около 7 649 Da и заметной тепловой стабильностью. IGF-2, с другой стороны, является одноцепным легко кислым белком с 67 аминокислотами и молекулярной массой около 7 471 Da, и стабильен в 0,1% SDS. 2 разделяют более 70% гомологии последовательности и примерно 50% структурного и функционального сходства с человеческим проинсулином.
-
Трансформационные факторы роста (TGF)
Факторы роста трансформации (TGF) относятся к двум классам факторов роста полипептидов: TGF-α и TGF-β. TGF-α производится макрофагами, клетками мозга и эпидермальными клетками и играет роль в индукции развития эпителия.
У человека TGF-β существует в трех изоформах: TGF-β1, TGF-β2 и TGF-β3. Эти изоформы связываются с общим рецепторным комплексом, состоящим из рецепторов TGF-β типа I (TGF-βR1) и типа II (TGF-βR2), индуцируя аналогичные внутриклеточные сигнальные каскады in vitro [6].
TGF-β (Transforming Growth Factor Beta) является многофункциональным белком, который регулирует широкий спектр клеточных процессов, включая рост клеток, дифференциацию, апоптоз и иммунную модуляцию. Рецепторы TGF-β представляют собой рецепторы серин/треонинкиназы, и сигнализация нижнего потока в основном посредничествует через путь SMAD и/или путь DAXX.
-
Фактор роста соединительной ткани (CTGF)
Фактор роста соединительной ткани (CTGF), также известный как FISP12 или CCN2, принадлежит к семейству белков CCN, которое включает в себя 3 члена: Cyr61, CTGF и Nov. Эти члены семьи демонстрируют высокую степень гомологии аминокислотных последовательностей и разделяют консервированную структуру белка, включающую четыре ключевых домена:
- N-терминальный инсулиноподобный фактор роста (IGFBP),
- повторение фактора фон-Виллебранда типа С (VWC),
- повторение тромбоспондина типа 1 (TSP1),
- и богатый цистеином С-терминальный домен (КТ).
CTGF был впервые идентифицирован в 1991 году как фибробластно-индуцируемый секретируемый белок-12 (FISP12), изолированный из клеток NIH3T3, активированных сывороткой. Название «фактор роста соединительной ткани» было предложено позже в том же году.
CTGF первоначально был признан своей способностью способствовать пролиферации фибробластов, миграции, адгезии и производству внеклеточной матрицы (ECM). Каждый из его структурных доменов может связывать специфические белки-партнеры, что позволяет CTGF выполнять широкий спектр биологических функций [7] (см. рисунок 3).
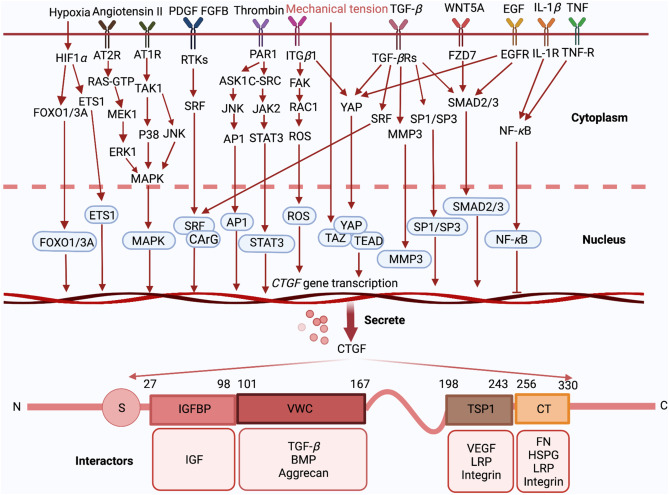
Рисунок 3
При внешней стимуляции активируются сигнальные пути, связанные с CTGF, что приводит к транскрипции и секреции CTGF. После секреции четыре отдельных домена CTGF - IGFBP, VWC, TSP1 и CT - могут взаимодействовать с различными молекулами для выполнения своих физиологических функций.
-
Фактор роста кератиноцитов (KGF)
Фактор роста кератиноцитов (KGF) является основным белковым фактором роста, секретируемым мезенхимальными клетками в подкожных тканях. Он конкретно стимулирует различные физиологические процессы в эпителиальных клетках, включая метаболизм, регенерацию, дифференциацию и миграцию. КГФ - это природный, биоактивный, растворимый белок в организме человека, кодированный последовательностью 194 аминокислот. Зрелая форма КГФ состоит из 163 аминокислотных остатков и содержит N-терминальное место гликозилации.
КГФ в основном секретируется подкожной тканью и связывается конкретно с его рецептором на поверхности эпителиальных клеток. Через сложный сигнальный каскад он активирует экспрессию генов, участвующих в делении и росте клеток, стимулируя тем самым метаболическую активность и регенерацию эпителиальных тканей.
-
Фактор роста нервов (NGF)
Семейство факторов роста нервов (NGF) включает в себя несколько нейротрофических факторов: сам NGF, нейротрофический фактор мозга (BDNF), нейротрофин-3 (NT-3), нейротрофин-4/5 (NT-4/5) и, в меньшей степени, нейротрофин-6 и нейротрофин-7. Среди них первые четыре являются наиболее известными и хорошо изученными.
NGF является наиболее важным и представительным членом этой семьи. В тканях НГФ в основном присутствует в форме прекурсора, который перерабатывается в зрелую форму в основном в субмандибулярной железе. NGF играет множество ролей, включая поддержку выживания нейронов, защиту нервов, содействие регенерации нервов, модулирование иммунных ответов, облегчение заживления ран и ингибирование роста опухоли.
В дополнение к вышеупомянутым факторам роста, другие важные члены включают факторы роста, связанные с интерлейкином (такие как ИЛ-1 и ИЛ-3), эритропоетин (ЭПО) и факторы, стимулирующие колонии (КСФ). Эти молекулы также играют решающую роль в регулировании пролиферации клеток, дифференциации и иммунных ответов в различных физиологических и патологических контекстах.
Механизм действий
-
Способы действия
Эндокринные факторы роста секретируются и транспортируются через кровоток, чтобы действовать на удаленные клетки-мишень. Примером является фактор роста тромбоцитов (PDGF).
Факторы роста, секретируемые клеткой, действуют на близлежащие клетки другого типа, влияя на их поведение.
Автокринный: факторы роста действуют на ту же клетку, которая их синтезировала и секретировала.
Среди них паракринная и автокринная сигнализация являются преобладающими способами действия большинства факторов роста.
-
Механический путь
После секретирования различными типами клеток факторы роста (ГФ) оказывают свое биологическое воздействие, связываясь с конкретными рецепторами, расположенными на поверхности или внутри клеток-мишеней. При связывании лиганда эти рецепторы активируют внутриклеточные сигнальные каскады, которые посредничествуют в соответствующих биологических ответах.
Основный класс рецепторов фактора роста обладает внутренней активностью тирозинкиназы и известен как рецепторные тирозинкиназы (RTK). RTK делятся примерно на 20 семейств, включая семейство рецепторов эпидермального фактора роста (EGFR), семейство рецепторов фактора роста тромбоцитов (PDGFR) и семейство рецепторов фактора роста нервов (NGFR). Действие большинства факторов роста опосредовано через РТК (см. рисунок 4).
Когда фактор роста связывается со своим рецептором на поверхности клетки, трансдукция сигнала инициируется димеризацией RTK и последующей активацией их киназных доменов. Два мономера внутри димера RTK подвергаются перекрестной фосфорилации на остатках тирозина, полностью активируя рецептор. Эти остатки фосфорилированного тирозина на С-терминальных хвостах RTK служат местами стыковки для нескольких молекул сигнализации ниже потока.
Формирование этих RTK сигнальных комплексов приводит к активации различных потоковых сигнальных путей, включая Ras/Erk, PI3K/Akt, Src/Jak/Stat и PLC-γ1. Эти пути взаимодействуют друг с другом, образуя сложную сигнальную сеть [8].
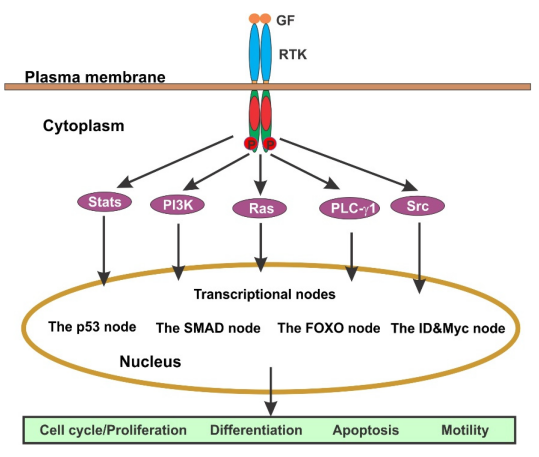
Рисунок 4
RTK-Ras сигнальный путь:
Связывание лиганда → Активация рецепторной тирозинкиназы (RTK) → Активированный RTK набирает адапторные белки → Фактор обмена нуклеотидами гуанина (ГЭФ) способствует высвобождению ВВП → Активация Ras (GTP-связывающего белка).
Эта активация вызывает каскад нижеследующих событий:
Активированный Ras инициирует активацию Raf, сериновой/треониновой белковой киназы (также известной как MAPKKK), которая фосфорилирует остатки серина/треонина на целевых белках. → Активированный Raf связывается и фосфорилирует MAPKK (MAPK киназа), приводя к его активации. MAPKK является киназой двойной специфичности, способной фосфорилировать остатки треонина и тирозина на MAPK, тем самым активируя MAPK. → Активированная MAPK транслокируется в ядро → Фосфорилирование других киназ или регулирующих гены белков (т.е. факторов транскрипции), модулирующих экспрессию генов (см. рисунок 5).
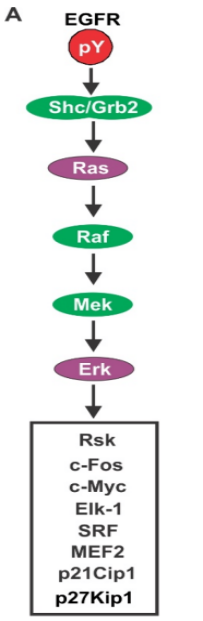
Рисунок 5
Ссылки
Каджи К. Функция, молекулярная структура и регулирование экспрессии генов фактора роста тромбоцитов. Нихон Риншо. август 1992 года; 50(8):1902-1909.
Heldin CH, Westermark B. Механизм действия и роль in vivo фактора роста тромбоцитов. Physiol Rev. 1999 октябрь; 79(4):1283-1316.
Такахаси С. Фактор роста сосудистой эндотелии (VEGF), рецепторы VEGF и их ингибиторы для антиангиогенной опухолевой терапии. Биол Фарм Булл. 2011; 34(12):1785-1788.
Ogiso H, Ishitani R, Nureki O, et al. Кристаллическая структура комплекса эпидермального фактора роста человека и рецепторных внеклеточных доменов. Клетка. 2002 Sep 20; 110(6):775-787.
Mossahebi-Mohammadi M, Quan M, Zhang JS, et al. FGF Signaling Pathway: A Key Regulator of Stem Cell Pluripotency (Сигнальный путь FGF: ключевой регулятор плюрипотенции стволовых клеток). Передняя клетка Dev Biol. 2020 Feb 18; 8:79.
Sun T, Vander Heiden JA, Gao X, et al. Изоформно-селективное ингибирование TGF-β3 для системного склероза. Med. 2024 9 февраля; 5(2):132-147.e7.
Fu M, Peng D, Lan T, et al. Многофункциональный регулирующий белок фактор роста соединительной ткани (CTGF): потенциальная терапевтическая цель для различных заболеваний. Acta Pharm Sin B. 2022 апрель; 12(4):1740-1760.
Ван З. Регулирование прогрессии клеточного цикла путем сигнализации клеток, индуцированной фактором роста. Cells 2021, 10, 3327.
Связанные продукты:
Если вы хотите получить более подробную информацию, пожалуйста, перейдите на наш сайт Магазина: www.solarbio.com.
|
Кошка нет. |
Название продукта |
Кошка нет. |
Название продукта |
|
СЕХ-0052 |
Человеческий VEGF ELISA Kit |
СЕХ-0050 |
Комплект ELISA для EGF человека |
|
СЕХ-0316 |
Человеческий набор ELISA TGF-β1 |
СЕКМ-0035 |
Набор ELISA TGF-β1 мыши |
|
СЕКМ-0039 |
Набор ELISA для мыши VEGF |
SEKR-0032 |
Набор ELISA VEGF для крыс |
|
SEKR-0010 |
Набор ELISA EGF для крыс |
SEKR-0012 |
Набор ELISA TGF-β1 для крыс |
 Консультируйтесь
Консультируйтесь
 в WhatsApp
в WhatsApp
 Оставить сообщение
Оставить сообщение
 верхний
верхний



