




ข่าวที่เกี่ยวข้อง


กลไกระดับโมเลกุลและเส้นทางการส่งสัญญาณของภาวะเครียดออกซิเดชัน: อะไรเกิดขึ้นจริงภายในเซลล์
ตารางเนื้อหา
ภาวะเครียดจากออกซิเดชันเป็นประเด็นวิจัยสำคัญในสาขาชีววิทยาของเซลล์ พิษวิทยา เภสัชวิทยา วิทยาศาสตร์ผู้สูงอายุ เนื้องอกวิทยา และการสร้างแบบจำลองโรคทางระบบประสาทเสื่อม คำจำกัดความพื้นฐานฟังดูตรงไปตรงมา: สารอนุมูลอิสระ (ROS) สะสมภายในเซลล์จนถึงระดับที่เกินความสามารถในการกำจัดสารต้านอนุมูลอิสระภายในเซลล์ และทำให้สมดุลของปฏิกิริยาออกซิเดชัน-รีดักชันเสียไป อย่างไรก็ตาม การทดลองเพาะเลี้ยงเซลล์ในทางปฏิบัติเผยให้เห็นตรรกะการควบคุมที่ซับซ้อนกว่ามาก การเพิ่มขึ้นของ ROS ในระดับเล็กน้อยและชั่วคราวทำหน้าที่เป็นตัวส่งสัญญาณรองที่สำคัญในการส่งสัญญาณทางสรีรวิทยา ในขณะที่การสะสม ROS ในระดับสูงอย่างต่อเนื่องจะก่อให้เกิดความเสียหายที่ไม่สามารถแก้ไขได้ต่อ DNA โปรตีน และไขมันในเยื่อหุ้มเซลล์ ส่งผลให้เกิดความผิดปกติของไมโทคอนเดรีย การอักเสบ การตายของเซลล์แบบอะพอพโทซิส และการตอบสนองต่อความเครียดของเซลล์ที่หลากหลาย
สำหรับห้องปฏิบัติการที่ทำการตรวจวัด ROS ด้วยการเรืองแสง การคัดกรองประสิทธิภาพสารต้านอนุมูลอิสระ และการวิจัยเกี่ยวกับกลไกการเกิดการอักเสบ การหาปริมาณ ขนาด ระยะเวลา และตำแหน่งภายในเซลล์ของการเพิ่มขึ้นของ ROS มีความสำคัญมากกว่าการยืนยันการเพิ่มขึ้นของ ROS ทั่วไปเพียงอย่างเดียว ตัวแปรแทรกซ้อนหลายอย่างสามารถทำให้ผลการทดลองคลาดเคลื่อนอย่างมาก รวมถึงแหล่งที่มาของเซลล์ ระยะเวลาการบ่มยา ประสิทธิภาพการโหลดโพรบเรืองแสง เกรดของสารเคมี โปรโตคอลการเก็บรักษาในอุณหภูมิต่ำ และการประมวลผลตัวอย่างหลังการรักษา โซลาร์บิโอ จัดจำหน่ายอุปกรณ์วิจัยด้านวิทยาศาสตร์ชีวภาพแบบครบวงจร ครอบคลุมถึงสารเคมีชีวภาพ สารประกอบชีวแอคทีฟโมเลกุลขนาดเล็ก แอนติบอดีปฐมภูมิ/ทุติยภูมิ ชุดตรวจ ELISA และวัสดุสิ้นเปลืองสำหรับการเพาะเลี้ยงเซลล์ ตอบสนองความต้องการด้านการวิจัยทางโมเลกุล ภูมิคุ้มกันวิทยา และชีวเคมีอย่างครบถ้วน และช่วยให้นักวิจัยลดอคติในการทดลองให้น้อยที่สุด
ภาวะเครียดจากออกซิเดชันคืออะไร?
ภาวะเครียดจากออกซิเดชัน หมายถึง สภาวะสมดุลของปฏิกิริยาออกซิเดชัน-รีดักชันภายในเซลล์ที่ถูกรบกวน ซึ่งเกิดจากการสร้าง ROS มากเกินไป หรือความสามารถในการกำจัดสารพิษโดยสารต้านอนุมูลอิสระบกพร่อง ในขณะที่ระดับ ROS ที่สูงอย่างต่อเนื่องก่อให้เกิดความเป็นพิษต่อเซลล์อย่างรุนแรง แต่ ROS ในปริมาณน้อยและเกิดขึ้นชั่วคราวทำหน้าที่เป็นตัวกลางในการส่งสัญญาณที่สำคัญในการปรับการทำงานของระบบภูมิคุ้มกัน การเพิ่มจำนวนเซลล์ และกระบวนการพัฒนาของเนื้อเยื่อ การเกิดภาวะเครียดจากออกซิเดชันถูกกระตุ้นโดยเงื่อนไขสำคัญสามประการ ได้แก่ การสะสม ROS ในปริมาณมากเกินไป การมี ROS มากเกินไปเป็นเวลานานภายในส่วนประกอบย่อยของเซลล์ที่ไวต่อปฏิกิริยาออกซิเดชัน-รีดักชัน (ไมโทคอนเดรียและนิวเคลียส) และสภาวะสมดุลของปฏิกิริยาออกซิเดชัน-รีดักชันที่ไม่สมดุลในระยะยาว ซึ่งทั้งหมดนี้จะเริ่มต้นกระบวนการที่เป็นพิษต่อเซลล์ในลำดับถัดไป
ประเภท ROS หลัก
ภายในเซลล์ยูคาริโอติกมีอนุมูลอิสระออกซิเจนหลากหลายชนิด โดยสามชนิดที่ได้รับการศึกษาอย่างกว้างขวางที่สุด ได้แก่ อนุมูลซูเปอร์ออกไซด์แอนไอออน (O2–•), ไฮโดรเจนเปอร์ออกไซด์ (ชม2O2) และอนุมูลไฮดรอกซิล (•โอ้)อนุมูลซูเปอร์ออกไซด์แอนไอออนส่วนใหญ่เกิดขึ้นควบคู่ไปกับกระบวนการขนส่งอิเล็กตรอนในไมโทคอนเดรีย ชม2O2 อนุมูลอิสระมีเสถียรภาพทางเคมีสูงและสามารถซึมผ่านเยื่อหุ้มเซลล์ได้ดี ทำให้สามารถเคลื่อนที่ข้ามขอบเขตของออร์แกเนลล์และทำหน้าที่เป็นทั้งสารส่งสัญญาณรีดอกซ์และตัวตรวจจับความเครียดของเซลล์ ในทางตรงกันข้าม อนุมูลไฮดรอกซิลมีปฏิกิริยาทางเคมีสูงมากและจะออกซิไดซ์ดีเอ็นเอ โปรตีน และไขมันในเยื่อหุ้มเซลล์ที่อยู่ใกล้เคียงทันทีที่เกิดขึ้น
การตรวจพบระดับ ROS รวมที่สูงขึ้นเพียงอย่างเดียวไม่เพียงพอที่จะสนับสนุนการวิจัยภาวะเครียดจากออกซิเดชันอย่างเข้มงวด การเพิ่มขึ้นของ ROS อาจเกิดจากสิ่งกระตุ้นเซลล์ที่แตกต่างกันโดยมีระยะเวลาที่แปรผัน และลำดับการส่งสัญญาณในขั้นตอนถัดไปแสดงให้เห็นถึงช่วงเวลาการกระตุ้นที่เหลื่อมกัน เพื่อให้ได้ข้อมูลการทดลองที่ครอบคลุมและสามารถทำซ้ำได้ นักวิจัยจำเป็นต้องมีชุดเครื่องมือวิจัยที่สมบูรณ์ ซึ่งครอบคลุมถึงแบบจำลองความเครียดของเซลล์ที่เสถียร ชุดตรวจจับทางชีวเคมีแบบหลายดัชนี และรีเอเจนต์สำหรับการวิเคราะห์เฉพาะเส้นทาง ซึ่งทั้งหมดนี้มีให้ครบถ้วนโดย โซลาร์บิโอ เพื่อลดความซับซ้อนของการวิเคราะห์โปรไฟล์รีดอกซ์แบบหลายมิติของคุณ
แหล่งกำเนิด ROS ภายในและภายนอกเซลล์
ไมโตคอนเดรียเป็นแหล่งกำเนิด ROS ภายในเซลล์ที่สำคัญที่สุด ในระหว่างกระบวนการออกซิเดชั่นฟอสโฟรีเลชั่นของไมโตคอนเดรีย การรั่วไหลของอิเล็กตรอนจากคอมเพล็กซ์ I และคอมเพล็กซ์ III ของห่วงโซ่การขนส่งอิเล็กตรอนทำให้อิเล็กตรอนอิสระทำปฏิกิริยากับออกซิเจนโมเลกุลเพื่อสร้างอนุมูลซูเปอร์ออกไซด์ เอนโดพลาสมิกเรติคูลัมยังสร้าง ROS ภายใต้ภาวะเครียดจากการตอบสนองต่อโปรตีนที่พับตัวผิดปกติ การออกซิเดชั่นเบต้าของกรดไขมันในเพอร์ออกซิโซมและการกระตุ้น NADPH oxidase (แหล่งกำเนิด ROS หลักในแบบจำลองเซลล์ภูมิคุ้มกันและเซลล์อักเสบ) ยังช่วยเพิ่มปริมาณ ROS ภายในเซลล์อีกด้วย
ปัจจัยภายนอกที่ก่อให้เกิดความเครียดสามารถเพิ่มปริมาณ ROS ในเซลล์ได้อย่างมาก เช่น รังสีอัลตราไวโอเลตและรังสีไอออนไนซ์กระตุ้นการสร้างอนุมูลอิสระโดยตรง โลหะหนักยับยั้งการทำงานของเอนไซม์ต้านอนุมูลอิสระและรบกวนการทำงานของไมโทคอนเดรีย สารพิษที่เกี่ยวข้องกับยาสูบ เอทานอลในปริมาณมาก ฝุ่นละอองในอากาศ และสารเคมีบำบัด ล้วนทำลายสมดุลของปฏิกิริยาออกซิเดชัน-รีดักชัน ที่สำคัญคือ สภาวะการเพาะเลี้ยงเซลล์ตามปกติสามารถเปลี่ยนแปลงระดับ ROS พื้นฐานและทำให้เกิดความคลาดเคลื่อนในการทดลองได้ เช่น สูตรอาหารเลี้ยงเซลล์ ลำดับของซีรั่มจากลูกวัว จำนวนรอบการเพาะเลี้ยงเซลล์ และความหนาแน่นของการเพาะเลี้ยง ตัวแปรที่มองข้ามได้ง่ายเหล่านี้มักนำไปสู่ความคลาดเคลื่อนอย่างมากระหว่างกลุ่มทดลองคู่ขนาน โซลาร์บิโอ จัดเตรียมอาหารเลี้ยงเซลล์มาตรฐาน เซรั่มที่มีเอนโดท็อกซินต่ำ และสารแยกเซลล์ที่มีความสม่ำเสมอ เพื่อลดการรบกวนในการทดลองให้น้อยที่สุด
ROS ทำลายเซลล์ได้อย่างไร?
โดยปกติแล้ว ROS ทำหน้าที่เป็นโมเลกุลส่งสัญญาณ อย่างไรก็ตาม หากมีปริมาณมากเกินไป มันอาจเริ่มทำลายส่วนประกอบสำคัญของเซลล์ได้
ความเสียหายของดีเอ็นเอ
อนุมูลไฮดรอกซิลที่มีปฏิกิริยาสูงจะกระตุ้นการเปลี่ยนแปลงออกซิเดชันของเบสกัวนีนในดีเอ็นเอ ทำให้เกิด 8-OHdG ซึ่งเป็นตัวบ่งชี้ทางชีวภาพหลักของความเสียหายจากออกซิเดชันต่อดีเอ็นเอ อนุมูลเหล่านี้ยังทำให้เกิดการแตกหักของดีเอ็นเอทั้งแบบสายเดี่ยวและสายคู่ และการเชื่อมโยงข้ามระหว่างดีเอ็นเอและโปรตีนที่ไม่สามารถย้อนกลับได้ ความเสียหายของดีเอ็นเอชั่วคราวจะกระตุ้นให้เซลล์หยุดวงจรชั่วคราวและกระตุ้นกลไกการซ่อมแซมดีเอ็นเอภายในร่างกาย แต่ความเสียหายจากออกซิเดชันที่เกิดขึ้นอย่างต่อเนื่องหรือซ้ำๆ จะนำไปสู่ความไม่เสถียรของจีโนม การเสื่อมสภาพของเซลล์ และการเปลี่ยนแปลงไปเป็นเซลล์มะเร็ง
การออกซิเดชันของโปรตีน
ROS จะออกซิไดซ์หมู่ข้างเคียงของกรดอะมิโนภายในโพลีเปปไทด์โดยตรง: หมู่ซัลฟ์ไฮดริลของซิสเทอีนจะเกิดการออกซิเดชันแบบย้อนกลับได้ ในขณะที่หมู่เมทิลีนของเมไทโอนีนจะสร้างพันธะคู่คาร์บอน-คาร์บอนหลังจากการออกซิเดชัน ความเข้มข้นของการออกซิเดชันส่งผลให้เกิดผลลัพธ์ทางหน้าที่ที่ตรงกันข้ามกันอย่างสิ้นเชิง: การออกซิเดชันของซิสเทอีนในระดับอ่อนทำหน้าที่เป็นสวิตช์รีดอกซ์แบบย้อนกลับได้สำหรับการส่งสัญญาณ ในขณะที่การดัดแปลงออกซิเดชันที่รุนแรงจะกระตุ้นให้เกิดการพับตัวของโปรตีนผิดปกติอย่างถาวร การไม่ทำงานของฟังก์ชัน และการก่อตัวของสารรวมตัวที่ไม่ละลายน้ำ เส้นทางนี้เป็นหัวใจสำคัญของการวิจัยเกี่ยวกับความเสื่อมของระบบประสาท: การสะสม ROS มากเกินไปอย่างต่อเนื่องจะทำให้ระบบกำจัดโปรตีนที่เสียหายบกพร่องและเร่งการรวมตัวของคราบเบต้าอะไมลอยด์ ซึ่งเป็นลักษณะทางพยาธิวิทยาที่สำคัญของโรคอัลไซเมอร์
การเกิดออกซิเดชันของไขมัน
เยื่อหุ้มพลาสมาและออร์แกเนลล์อุดมไปด้วยกรดไขมันไม่อิ่มตัวเชิงซ้อน ซึ่งไวต่อการโจมตีจากอนุมูลอิสระ (ROS) อย่างมาก การเกิดปฏิกิริยาออกซิเดชันของไขมันก่อให้เกิดผลิตภัณฑ์สุดท้ายที่เสถียร ได้แก่ มาลอนไดอัลดีไฮด์ (MDA) และ 4-ไฮดรอกซีโนเนนอล (4-HNE) ซึ่งเป็นตัวบ่งชี้ทางชีวภาพที่วัดได้บ่อยที่สุดของการบาดเจ็บจากออกซิเดชันของไขมัน อัลดีไฮด์ที่เป็นพิษเหล่านี้ทำลายความสมบูรณ์ของเยื่อหุ้มเซลล์ ทำให้การทำงานของไมโทคอนเดรียบกพร่อง จับกับโปรตีนและดีเอ็นเอในเซลล์ และสร้างวงจรป้อนกลับเชิงบวกเพื่อขยายสัญญาณการอักเสบ เพื่อให้ได้ข้อมูลการทดลองที่แข็งแกร่งและครอบคลุมหลายมิติ กลุ่มวิจัยส่วนใหญ่จึงตรวจวัดปริมาณอนุมูลอิสระ (ROS) ระดับการเกิดปฏิกิริยาออกซิเดชันของไขมัน กิจกรรมของเอนไซม์ต้านอนุมูลอิสระ การหลั่งไซโตไคน์ที่ก่อให้เกิดการอักเสบ และความมีชีวิตของเซลล์ไปพร้อมกัน
กลไกการส่งสัญญาณที่สำคัญในภาวะเครียดออกซิเดชัน
ภาวะเครียดจากออกซิเดชันไม่ได้ส่งผลให้เกิดกระบวนการทางเซลล์เพียงอย่างเดียว เนื่องจากอนุมูลอิสระ (ROS) สามารถทำลายโมเลกุลแต่ละตัวได้ รวมถึงกระตุ้นหรือปรับเปลี่ยนกระบวนการต่างๆ ได้ด้วย เส้นทางการส่งสัญญาณภาวะเครียดจากออกซิเดชันสามารถกระตุ้นให้เกิดการตอบสนองของเซลล์ได้หลากหลายรูปแบบ ซึ่งขึ้นอยู่กับความรุนแรงและระยะเวลาของภาวะเครียดจากออกซิเดชัน รวมถึงสภาวะพื้นฐานของเซลล์ด้วย
วิถีการทำงานของ Nrf2-ARE: การป้องกันสารต้านอนุมูลอิสระ
กลไกการทำงานของ Nrf2-ARE ทำหน้าที่เป็นเส้นทางควบคุมหลักในการป้องกันสารต้านอนุมูลอิสระในเซลล์ ภายใต้สภาวะสมดุลของปฏิกิริยาออกซิเดชัน-รีดักชันในสภาวะปกติ Nrf2 ในไซโตพลาสซึมจะจับกับ Keap1 อย่างแน่นหนาและ undergoes การยูบิควิตินเนชันและการย่อยสลายโดยโปรตีเอโซมอย่างต่อเนื่อง เมื่อ ROS สะสมมากขึ้น อนุมูลอิสระจะดัดแปลงหมู่ซิสเทอีนบน Keap1 ด้วยพันธะโควาเลนต์ ทำให้โครงสร้างของ Keap1 เปลี่ยนไปและทำให้ Nrf2 แยกตัวออก ในขณะเดียวกัน PI3K/Akt ที่ถูกกระตุ้นจะฟอสโฟรีเลต Nrf2 เพื่อเร่งการเคลื่อนย้ายไปยังนิวเคลียสให้เร็วขึ้น ภายในนิวเคลียส Nrf2 จะสร้างเฮเทอโรไดเมอร์กับโปรตีน Maf ขนาดเล็กและจับกับองค์ประกอบตอบสนองต่อสารต้านอนุมูลอิสระ (AREs) บนโปรโมเตอร์ของยีนเป้าหมาย
โปรแกรมการถอดรหัสทางพันธุกรรมนี้จะเพิ่มการทำงานของเอนไซม์ต้านอนุมูลอิสระ (SOD, CAT, GSH-Px) เอนไซม์สังเคราะห์กลูตาไธโอน และโปรตีนล้างพิษระยะที่สอง ซึ่งโดยรวมแล้วจะช่วยเพิ่มความสามารถในการกำจัด ROS และซ่อมแซมความเสียหายของเซลล์ แบบจำลองทางคลินิกก่อนหน้านี้จำนวนมากยืนยันว่าการกระตุ้น Nrf2 มีผลในการปกป้องเซลล์ในเนื้อเยื่อตับ ปอด ระบบประสาท และเนื้อเยื่อบุผิวที่ได้รับความเสียหาย
ที่น่าสังเกตคือ การอ่านค่าของวิถีการทำงานแสดงให้เห็นถึงพลวัตเชิงเวลาที่ไม่สอดคล้องกัน: การเคลื่อนย้ายของ Nrf2 ไปยังนิวเคลียส การแสดงออกของ mRNA/โปรตีนของยีนเป้าหมายปลายทาง กิจกรรมของเอนไซม์ต้านอนุมูลอิสระ และประสิทธิภาพการกำจัด ROS ไม่ได้ถึงค่าสูงสุดพร้อมกัน
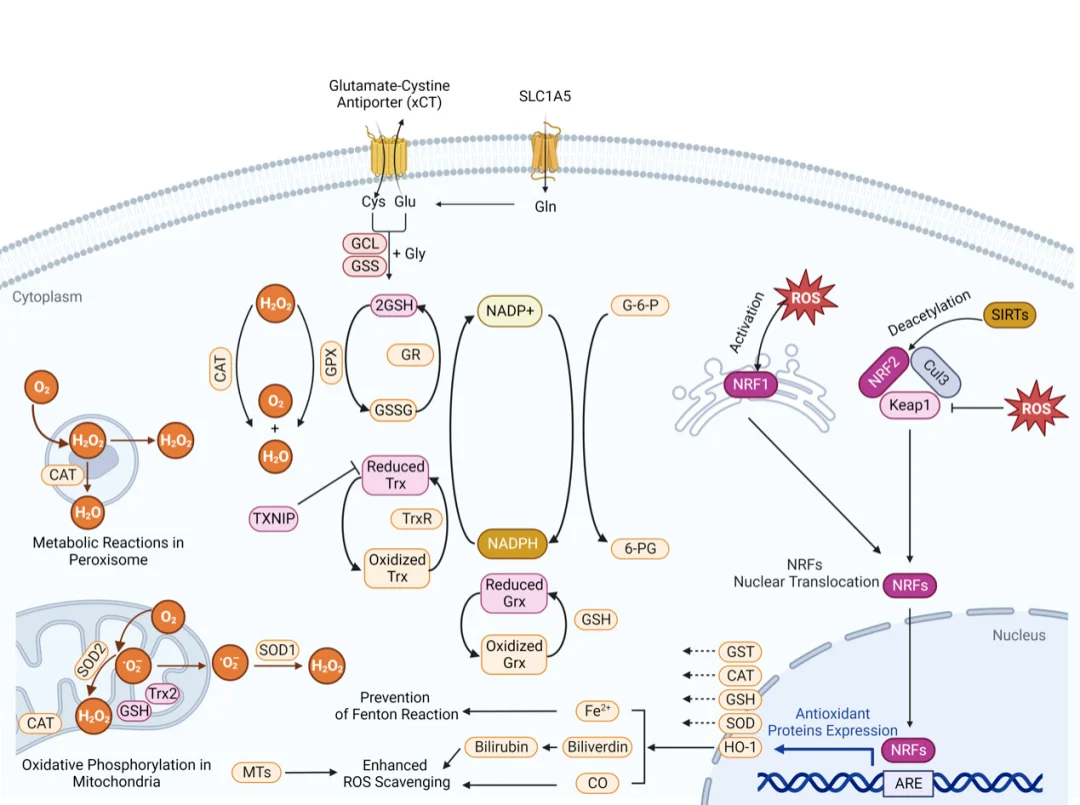
วิถี NF-κB: ศูนย์กลางการสื่อสารระหว่างภาวะเครียดออกซิเดชันและการอักเสบ
กลไกการทำงานของ NF-κB ทำหน้าที่เป็นสะพานโมเลกุลที่สำคัญในการเชื่อมโยงระหว่างภาวะ ROS เกินและการตอบสนองต่อการอักเสบ ในเซลล์ที่อยู่ในสภาวะพักและไม่ได้รับการกระตุ้น ไดเมอร์ของ NF-κB จะจับกับโปรตีน IκB ที่ยับยั้งการทำงานอย่างแน่นหนาและถูกกักเก็บไว้ภายในไซโตพลาสซึม การเพิ่มขึ้นของ ROS จะกระตุ้นคอมเพล็กซ์ไคเนส IKK ซึ่งจะฟอสโฟรีเลต IκB เพื่อกระตุ้นการยูบิควิตินเนชันและการย่อยสลายโดยโปรตีเอโซม NF-κB ที่เป็นอิสระจะเคลื่อนย้ายไปยังนิวเคลียสและกระตุ้นการถอดรหัสของสารสื่อกลางที่ก่อให้เกิดการอักเสบ ได้แก่ TNF-α, IL-1β และ IL-6
เส้นทางนี้ก่อให้เกิดวงจรป้อนกลับเชิงบวกที่ทวีความรุนแรงขึ้นเอง: ROS เริ่มต้นการส่งสัญญาณการอักเสบ และเซลล์ภูมิคุ้มกันที่ถูกกระตุ้นจะหลั่ง ROS เพิ่มเติมเพื่อเพิ่มกิจกรรมของ NF-κB ให้มากขึ้น การกระตุ้นแบบวนซ้ำอย่างต่อเนื่องนี้ขับเคลื่อนความก้าวหน้าของโรคอักเสบเรื้อรัง รวมถึงหลอดเลือดแดงแข็ง โรคข้ออักเสบรูมาตอยด์ การบาดเจ็บจากอนุมูลอิสระในตับ และการอักเสบเรื้อรังระดับต่ำในระบบเผาผลาญ
MAPK Cascade: ตัวควบคุมหลักของการเพิ่มจำนวนเซลล์ การตอบสนองต่อความเครียด และอะพอพโทซิส
วิถีการส่งสัญญาณของโปรตีนไคเนสที่กระตุ้นด้วยไมโทเจน (MAPK) ถูกกระตุ้นโดยปัจจัยความเครียดในเซลล์ที่หลากหลาย รวมถึง ROS ซึ่งประกอบด้วยสามกลุ่มย่อยที่มีหน้าที่แตกต่างกัน ได้แก่ ERK, JNK และ p38 MAPK ซึ่งทำงานผ่านการฟอสฟอริเลชันที่ควบคุมโดย MKK อย่างต่อเนื่อง ความเข้มข้นของ ROS ในระดับต่ำตามสภาวะทางสรีรวิทยาจะกระตุ้นการส่งสัญญาณของ ERK อย่างเลือกสรร เพื่อช่วยให้เซลล์ดำเนินไปในวงจรชีวิต ขยายพันธุ์ และรักษาสมดุลของปฏิกิริยาออกซิเดชัน-รีดักชัน ในทางตรงกันข้าม การสะสมของ ROS ที่มากเกินไปจะกระตุ้นการฟอสฟอริเลชันของ JNK และ p38 อย่างรุนแรง ทำให้เกิดการตอบสนองต่อความเครียดในเซลล์ การถอดรหัสยีนที่ก่อให้เกิดการอักเสบ และการตายของเซลล์แบบอะพอพโทซิส
ในแบบจำลองการบาดเจ็บจากภาวะขาดเลือดและฟื้นฟูการไหลเวียนโลหิตของกล้ามเนื้อหัวใจ การนำออกซิเจนกลับเข้าไปอย่างรวดเร็วจะกระตุ้นให้เกิด ROS ในปริมาณมาก ทำให้การส่งสัญญาณ JNK/p38 ทำงานมากเกินไปและกระตุ้นให้เกิดการตายของเซลล์กล้ามเนื้อหัวใจจำนวนมาก ในการวิจัยด้านมะเร็งวิทยา ROS ในระดับต่ำพื้นฐานจะช่วยส่งเสริมการเจริญเติบโตของเนื้องอกผ่านการกระตุ้น ERK ในขณะที่ระดับ ROS ที่เพิ่มขึ้นจากการใช้ยาจะทำให้เซลล์มะเร็งไวต่อเคมีบำบัดและรังสีบำบัดมากขึ้น โดยรวมแล้ว ทั้งความเข้มข้นของ ROS และระยะเวลาการสัมผัสจะเป็นตัวกำหนดชะตากรรมของเซลล์ในขั้นสุดท้าย ซึ่งถูกควบคุมโดยการส่งสัญญาณ MAPK
เส้นทาง PI3K/Akt: สัญญาณการอยู่รอดเพื่อปกป้องเซลล์ภายใต้ภาวะความเครียดจากปฏิกิริยาออกซิเดชัน-รีดักชันระดับปานกลาง
ระบบส่งสัญญาณ PI3K/Akt เป็นแกนส่งสัญญาณหลักที่ช่วยให้เซลล์อยู่รอดได้ภายใต้ภาวะเครียดออกซิเดชันระดับอ่อน การกระตุ้นด้วย ROS ในระดับปานกลางจะกระตุ้น PI3K ให้สร้าง PIP3 ที่ยึดติดกับเยื่อหุ้มเซลล์ ซึ่งจะดึงดูด Akt จากไซโตพลาสซึมไปยังเยื่อหุ้มเซลล์เพื่อทำการฟอสฟอริเลชันและกระตุ้นการทำงานอย่างเต็มที่ Akt ที่ถูกฟอสฟอริเลชันจะยับยั้งการส่งสัญญาณอะพอพโทซิสผ่านกลไกหลายอย่าง ได้แก่ การฟอสฟอริเลชันและยับยั้งโปรตีน Bad ซึ่งเป็นโปรตีนที่กระตุ้นอะพอพโทซิส ยับยั้งการทำงานของแคสเปส และเพิ่มการเผาผลาญและการเพิ่มจำนวนของเซลล์ที่ควบคุมโดย mTOR นอกจากนี้ Akt ยังฟอสฟอริเลชัน Nrf2 โดยตรงเพื่อเร่งการเคลื่อนย้ายไปยังนิวเคลียสและเพิ่มการตอบสนองการถอดรหัสของสารต้านอนุมูลอิสระ
การส่งสัญญาณป้องกันนี้มีเกณฑ์รีดอกซ์ที่สำคัญ เมื่อความเข้มข้นของ ROS เกินช่วงที่ยอมรับได้ PI3K และ Akt จะถูกทำลายโดยปฏิกิริยาออกซิเดชันและย่อยสลายด้วยเอนไซม์อย่างถาวร ทำให้การส่งสัญญาณภายในเซลล์เปลี่ยนไปสู่เส้นทางอะพอพโทซิสภายในไมโทคอนเดรีย ในแบบจำลองเซลล์ประสาทของโรคพาร์กินสัน ประสิทธิภาพในการรักษาด้วยสารต้านอนุมูลอิสระขึ้นอยู่กับสถานะการทำงานที่สมบูรณ์ของระบบส่งสัญญาณ PI3K/Akt เป็นอย่างมาก
ความเชื่อมโยงระหว่างโรคต่างๆ ในงานวิจัยเกี่ยวกับภาวะเครียดออกซิเดชัน
ภาวะเครียดจากออกซิเดชันเป็นตัวขับเคลื่อนทางพยาธิวิทยาที่พบได้ทั่วไปในความผิดปกติมากมายของมนุษย์ การวิเคราะห์ลำดับการส่งสัญญาณรีดอกซ์ที่อยู่เบื้องหลังซึ่งเป็นสาเหตุของการบาดเจ็บของเนื้อเยื่อ แทนที่จะระบุเพียงแค่ลักษณะความเสียหายของเซลล์นั้น จะให้คุณค่าในการวิจัยเชิงประยุกต์มากกว่ามาก
การวิจัยมะเร็ง
เซลล์มะเร็งมีสถานะ ROS พื้นฐานที่แตกต่างกัน: บางชนิดรักษาระดับ ROS สูงโดยธรรมชาติ ในขณะที่บางชนิดเพิ่มการทำงานของแกนต้านอนุมูลอิสระ Nrf2 เพื่อยับยั้งการสะสมของ ROS ซึ่งทำให้มีความต้านทานต่อเคมีบำบัดและรังสีบำบัดสูง อย่างไรก็ตาม การเพิ่มระดับ ROS ภายในเซลล์ให้สูงถึงระดับที่เป็นพิษสามารถกระตุ้นให้เซลล์มะเร็งตายได้สำเร็จ สำหรับการวิจัยด้านมะเร็งวิทยา ระบบตรวจจับแบบหลายมิติมีความสำคัญอย่างยิ่ง รวมถึงโพรบเรืองแสง ROS ชุดทดสอบความไวต่อยา คลังสารประกอบโมเลกุลขนาดเล็ก และแอนติบอดีไบโอมาร์กเกอร์เฉพาะเส้นทาง
การวิจัยโรคความเสื่อมของระบบประสาท
เซลล์ประสาทที่เจริญเต็มที่แล้วเป็นเซลล์ที่ไม่สามารถแบ่งตัวได้อีกต่อไป ไม่สามารถสร้างใหม่ได้ มีกระบวนการเผาผลาญพลังงานที่เปราะบาง และมีความอ่อนแออย่างมากต่ออันตรายจากอนุมูลอิสระ ความเสียหายที่เกิดจาก ROS ต่อไมโทคอนเดรียและระบบการพับโปรตีนจะนำไปสู่การตายของเซลล์แบบอะพอพโทซิสในที่สุด แบบจำลองสัตว์ทดลองของโรคพาร์กินสันแสดงให้เห็นว่า ROS ที่มากเกินไปจะยับยั้งวิถี PI3K/Akt ที่ช่วยปกป้องเซลล์ และกระตุ้นการส่งสัญญาณ JNK ที่ทำให้เกิดอะพอพโทซิสมากเกินไป ในโรคอัลไซเมอร์ การสะสมของ ROS มากเกินไปจะเร่งการสะสมของเบต้า-อะไมลอยด์ที่ผิดปกติ และกระตุ้นให้เกิดการอักเสบของระบบประสาทอย่างต่อเนื่อง การกระตุ้น Nrf2 อย่างจำเพาะเจาะจงสามารถบรรเทาอาการเสื่อมของระบบประสาทเหล่านี้ได้อย่างมีประสิทธิภาพ
แบบจำลองการบาดเจ็บของหัวใจและตับ
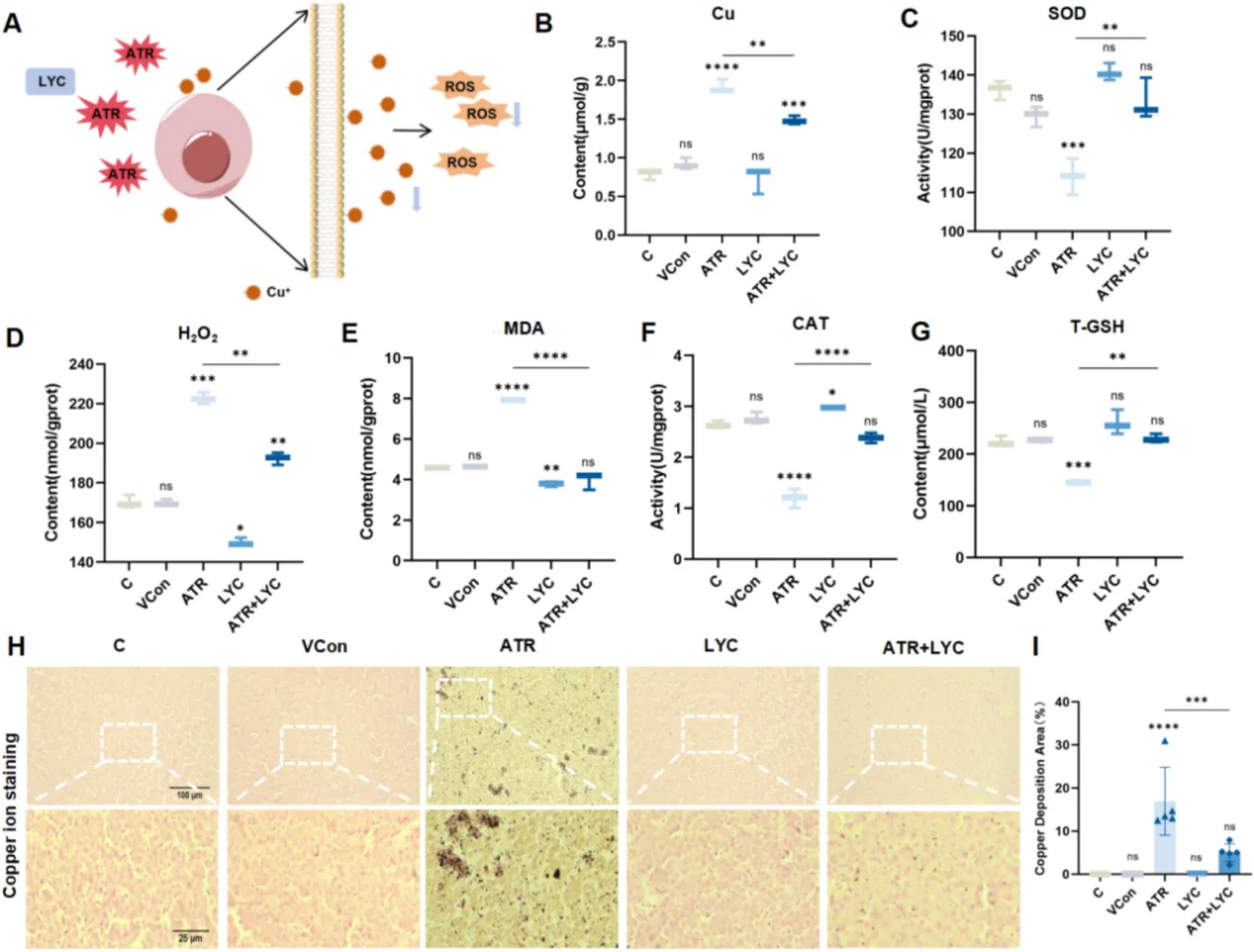
ในการวิจัยเกี่ยวกับระบบหัวใจและหลอดเลือด การกระตุ้น NF-κB อย่างต่อเนื่องจะทำให้เกิดการอักเสบของเยื่อบุหลอดเลือดและการแสดงออกของโมเลกุลยึดเกาะมากเกินไป ซึ่งเร่งการก่อตัวของคราบพลัคในหลอดเลือดแดง การระเบิดของ ROS อย่างรวดเร็วที่เกิดจากภาวะขาดเลือดและภาวะเลือดไหลเวียนกลับสู่กล้ามเนื้อหัวใจจะกระตุ้นการทำงานของ JNK/p38 MAPK และกระตุ้นให้เกิดการตายของเซลล์กล้ามเนื้อหัวใจจำนวนมาก สำหรับแบบจำลองพิษวิทยาของตับ ความเครียดจากออกซิเดชันจะกระตุ้นให้เกิดความผิดปกติของไมโทคอนเดรีย การเกิดปฏิกิริยาออกซิเดชันของไขมันจำนวนมาก การหลั่งไซโตไคน์ที่ก่อให้เกิดการอักเสบ และการตายของเซลล์ที่มีทองแดงเป็นองค์ประกอบ แบบจำลองการวิจัยทั่วไปใช้สารอะทราซีนเพื่อเหนี่ยวนำให้เกิดความเสียหายจากออกซิเดชันในตับ โดยใช้ไลโคปีนเป็นสารต้านอนุมูลอิสระเพื่อฟื้นฟูเนื้อเยื่อที่เสียหาย
คำแนะนำการใช้งานผลิตภัณฑ์สำหรับการทดลองเกี่ยวกับภาวะเครียดออกซิเดชัน
สำหรับการทดลองกู้ภัยสารต้านอนุมูลอิสระ เราขอแนะนำ N-Acetyl L-Cysteine (รหัสสินค้า IA0050) ซึ่งเป็นสารโมเลกุลขนาดเล็กที่ทำหน้าที่เป็นตัวปรับสมดุลรีดอกซ์แบบคลาสสิกสำหรับการวิจัย ROS NAC ช่วยเติมเต็มกลุ่มไทออลภายในเซลล์และเสริมสร้างระบบต้านอนุมูลอิสระที่ขึ้นอยู่กับกลูตาไธโอน ทำให้ผู้วิจัยสามารถตรวจสอบได้ว่าการเปลี่ยนแปลงลักษณะที่สังเกตได้นั้นเกิดจากความเครียดที่เกิดจาก ROS หรือผลกระทบที่ไม่เกี่ยวข้องอื่นๆ
ไลโคปีน (รหัสสินค้า IL0510) เป็นสารต้านอนุมูลอิสระที่เชื่อถือได้อีกชนิดหนึ่งสำหรับการทดสอบในเซลล์ในหลอดทดลองและแบบจำลองเนื้อเยื่อสัตว์ในร่างกาย โดยเฉพาะอย่างยิ่งในการวิจัยเกี่ยวกับการบาดเจ็บของตับและการเกิดออกซิเดชันของไขมัน การประเมินประสิทธิภาพการต้านอนุมูลอิสระอย่างครอบคลุมนั้นจำเป็นต้องมีการตรวจวัดหลายดัชนีในระดับความเข้มข้นที่แตกต่างกัน การรบกวนจากตัวทำละลาย ระดับ ROS กิจกรรมต้านอนุมูลอิสระภายในเซลล์ และความมีชีวิตของเซลล์ การละเลยตัวชี้วัดใด ๆ เหล่านี้จะนำไปสู่ข้อสรุปเชิงกลไกที่ไม่สมบูรณ์และลำเอียง
ก่อนเริ่มการทดสอบการตรวจจับความเครียดจากออกซิเดชัน นักวิจัยต้องตรวจสอบเงื่อนไขการจัดเก็บสารเคมี ความสามารถในการละลายของตัวทำละลาย การรับรองคุณภาพของชุดการผลิต และช่วงเวลาการตรวจจับที่ถูกต้อง การอ่านค่าจากการทดลองอาจผิดเพี้ยนได้ง่ายจากความเสื่อมสภาพจากแสง การแช่แข็งและละลายซ้ำๆ และขั้นตอนการสลายเซลล์ที่ไม่สม่ำเสมอ ข้อมูลเกี่ยวกับการตั้งค่าการทดสอบหรือผลิตภัณฑ์ที่เหมาะสมสามารถดูได้ที่ [ลิงก์ไปยังเอกสารประกอบ] หน้าเว็บบริการด้านเทคนิคของ Solarbio หรือโดยการเรียกดูข้อมูลที่เกี่ยวข้อง บทความทางเทคนิค.
ข้อสรุป
ภาวะเครียดจากออกซิเดชันไม่สามารถนิยามได้ง่ายๆ ว่าเป็นเพียงการสะสมของ ROS ที่มากเกินไปเท่านั้น มันเกิดขึ้นจากเครือข่ายปฏิสัมพันธ์ที่ซับซ้อน: การผลิต ROS ที่ไม่ได้รับการควบคุมจะเกินขีดความสามารถของระบบกำจัดสารต้านอนุมูลอิสระภายในร่างกาย ทำให้เกิดการเปลี่ยนแปลงที่ไม่สามารถย้อนกลับได้ของ DNA โปรตีน และไขมันในเยื่อหุ้มเซลล์ สารเมตาบอไลต์ของไขมันที่ถูกออกซิไดซ์จะยิ่งกระตุ้นกระบวนการอักเสบ ซึ่งจะยิ่งทำให้ความเสียหายจากออกซิเดชันรุนแรงขึ้น และในที่สุดก็ทำให้เซลล์ตายแบบอะพอพโทซิส
เซลล์ใช้กลไกการส่งสัญญาณหลักสี่อย่างเพื่อตอบสนองต่อความไม่สมดุลของปฏิกิริยาออกซิเดชัน-รีดักชัน ได้แก่ เส้นทาง Nrf2-ARE เริ่มต้นการป้องกันสารต้านอนุมูลอิสระโดยรวม เส้นทาง NF-κB เป็นตัวกลางในการตอบสนองการอักเสบที่เกิดจากความเครียดจากออกซิเดชัน เส้นทาง MAPK กำหนดการเพิ่มจำนวนเซลล์หรือการตายของเซลล์โดยขึ้นอยู่กับปริมาณ ROS และเส้นทาง PI3K/Akt รักษาการอยู่รอดของเซลล์ภายใต้การรบกวนของปฏิกิริยาออกซิเดชัน-รีดักชันในระดับเล็กน้อย
การตรวจวัดด้วยดัชนีเดียวไม่สามารถรองรับการวิจัยภาวะเครียดออกซิเดชันอย่างเข้มงวดได้ เพื่อให้ได้ข้อมูลที่ครอบคลุมและสามารถทำซ้ำได้ ขอแนะนำอย่างยิ่งให้ตรวจวัดตัวบ่งชี้หลายตัวพร้อมกัน ซึ่งรวมถึงความเข้มข้นของ ROS ทั้งหมด กิจกรรมของเอนไซม์ต้านอนุมูลอิสระ ระดับการเกิดออกซิเดชันของไขมัน MDA/4-HNE สถานะการฟอสโฟรีเลชันของโปรตีนส่งสัญญาณที่สำคัญ การหลั่งไซโตไคน์ที่ก่อให้เกิดการอักเสบ การทำงานของไมโทคอนเดรีย ไบโอมาร์กเกอร์ที่เกี่ยวข้องกับอะพอพโทซิส และความมีชีวิตของเซลล์ หากคุณต้องการผลิตภัณฑ์ที่ตรงกับความต้องการเฉพาะ โปรโตคอลการทดลองโดยละเอียด หรือการสนับสนุนทางเทคนิคแบบตัวต่อตัว โปรดติดต่อเรา ทีมบริการวิจัยมืออาชีพของ Solarbio.
คำถามที่พบบ่อย
คำถามที่ 1: ภาวะเครียดจากออกซิเดชันคืออะไรกันแน่?
A1: ภาวะเครียดออกซิเดชัน หมายถึง การรบกวนสมดุลรีดอกซ์ภายในเซลล์ ซึ่งเกิดจากการสร้าง ROS มากเกินไป หรือความสามารถในการกำจัดสารต้านอนุมูลอิสระบกพร่อง ความไม่สมดุลนี้ก่อให้เกิดความเสียหายต่อโมเลกุลชีวภาพ การส่งสัญญาณผิดปกติ การตอบสนองต่อการอักเสบ และการตายของเซลล์แบบอะพอพโทซิส หรือการรวมกันของอาการทางพยาธิวิทยาเหล่านี้
คำถามที่ 2: สารอนุมูลอิสระ (ROS) เป็นอันตรายต่อเซลล์โดยเนื้อแท้หรือไม่?
A2: ไม่ใช่ ROS ที่มีความเข้มข้นต่ำและเกิดขึ้นชั่วคราวทำหน้าที่เป็นสารสื่อสารทางสรีรวิทยาที่จำเป็น มีเพียงระดับ ROS ที่สูงอย่างต่อเนื่อง หรือการสะสม ROS มากเกินไปเฉพาะที่ภายในออร์แกเนลล์ที่ไวต่อปฏิกิริยาออกซิเดชัน-รีดักชัน (ไมโตคอนเดรีย นิวเคลียส) เท่านั้นที่จะก่อให้เกิดผลกระทบที่เป็นพิษต่อเซลล์
คำถามที่ 3: กลไกการส่งสัญญาณหลักใดที่มีบทบาทสำคัญในการป้องกันสารต้านอนุมูลอิสระภายในเซลล์?
A3: วิถีสัญญาณ Nrf2-ARE ทำหน้าที่เป็นแกนควบคุมสารต้านอนุมูลอิสระหลัก โดยจะเพิ่มการแสดงออกของยีน SOD, CAT, GSH-Px, เอนไซม์สังเคราะห์ที่ขึ้นอยู่กับกลูตาไธโอน และโปรตีนล้างพิษระยะที่สอง เพื่อฟื้นฟูสมดุลของปฏิกิริยาออกซิเดชัน-รีดักชัน
ไตรมาสที่ 4: เหตุใด NF-κB จึงเป็นเป้าหมายสำคัญในการวิจัยเกี่ยวกับภาวะเครียดออกซิเดชัน?
A4: NF-κB เป็นโมเลกุลสำคัญที่เชื่อมโยงระหว่างภาวะ ROS เกินและการอักเสบ มันสร้างวงจรป้อนกลับเชิงบวกที่ขยายตัวอย่างต่อเนื่อง: ROS กระตุ้น NF-κB เพื่อเหนี่ยวนำให้เกิดการอักเสบ และเซลล์ภูมิคุ้มกันที่ถูกกระตุ้นจะหลั่ง ROS เพิ่มเติมเพื่อรักษากลไกการกระตุ้นต่อไป ทำให้ความเสียหายของเนื้อเยื่อเรื้อรังแย่ลง
Q5: ข้อควรระวังที่สำคัญใดบ้างที่ควรคำนึงถึงเมื่อใช้ NAC ในการทดลองเกี่ยวกับ ROS
A5: นักวิจัยจำเป็นต้องปรับความเข้มข้นของ NAC ระยะเวลาการบ่ม และสูตรตัวทำละลายให้เหมาะสมล่วงหน้า และทำการทดสอบความมีชีวิตของเซลล์ควบคู่กันไปเพื่อขจัดผลกระทบจากสารพิษต่อเซลล์ การตรวจจับไบโอมาร์กเกอร์รีดอกซ์แบบหลายดัชนีร่วมกันเป็นสิ่งจำเป็นเพื่อให้ได้ข้อสรุปเชิงทดลองที่น่าเชื่อถือและมั่นคง



