




Nouvelles connexes


Mécanismes moléculaires et voies de signalisation du stress oxydatif : que se passe-t-il réellement à l’intérieur des cellules ?
Table des matières
Le stress oxydatif représente un axe de recherche majeur en biologie cellulaire, toxicologie, pharmacologie, gérontologie, oncologie et modélisation des maladies neurodégénératives. Sa définition de base semble simple : les espèces réactives de l’oxygène (ERO) intracellulaires s’accumulent à des niveaux dépassant la capacité d’élimination antioxydante intrinsèque de la cellule et perturbent l’homéostasie redox. Cependant, des expériences pratiques en culture cellulaire révèlent une logique de régulation bien plus complexe. De faibles pics transitoires d’ERO jouent un rôle essentiel de messagers secondaires dans la signalisation physiologique, tandis qu’une surcharge soutenue et importante d’ERO induit des dommages irréversibles à l’ADN, aux protéines et aux lipides membranaires, entraînant successivement un dysfonctionnement mitochondrial, des cascades inflammatoires, la mort cellulaire par apoptose et diverses réponses au stress cellulaire.
Pour les laboratoires effectuant la détection par fluorescence des ROS, le criblage de l'efficacité des antioxydants et la recherche sur les voies de signalisation du stress inflammatoire, la quantification de l'amplitude, de la durée et de la localisation subcellulaire de l'élévation des ROS est bien plus importante que la simple confirmation d'une augmentation générique de ces dernières. De multiples variables confondantes peuvent fausser considérablement les résultats expérimentaux, notamment l'origine de la lignée cellulaire, la durée d'incubation du médicament, l'efficacité du chargement de la sonde fluorescente, la qualité des réactifs, les protocoles de conservation à froid et le traitement des échantillons après application. Solarbio fournit des produits complets pour la recherche en sciences de la vie, notamment des réactifs biochimiques, des composés bioactifs à petites molécules, des anticorps primaires/secondaires, des kits de détection ELISA et des consommables pour la culture cellulaire, répondant pleinement aux besoins de la recherche sur les voies moléculaires, immunologiques et biochimiques et aidant les chercheurs à minimiser les biais expérimentaux.
Qu'est-ce que le stress oxydatif ?
Le stress oxydatif désigne une perturbation de l'équilibre redox intracellulaire, causée soit par une production excessive d'espèces réactives de l'oxygène (ROS), soit par une altération des capacités de détoxification antioxydante. Si des niveaux élevés et soutenus de ROS exercent une forte cytotoxicité, des concentrations transitoires et faibles de ROS jouent un rôle essentiel de médiateurs de signalisation, modulant l'activation immunitaire, la prolifération cellulaire et les processus de développement tissulaire. L'apparition du stress oxydatif est déclenchée par trois conditions clés : une accumulation excessive de ROS, une surcharge prolongée en ROS au sein des compartiments subcellulaires sensibles au redox (mitochondries et noyau) et un déséquilibre redox persistant, autant de facteurs qui initient des cascades cytotoxiques en aval.
Principaux types de ROS
Diverses espèces réactives de l'oxygène existent au sein des cellules eucaryotes ; les trois sous-types les plus étudiés sont l'anion radical superoxyde (O2–•), peroxyde d'hydrogène (H2O2), et le radical hydroxyle (•OH)Les radicaux anions superoxydes sont principalement générés le long des chaînes de transport d'électrons mitochondriales. H2O2 Elle présente une stabilité chimique supérieure et une perméabilité membranaire élevée, ce qui lui permet de traverser les frontières des organites et d'agir à la fois comme messager de signalisation redox et comme capteur de stress cellulaire. À l'inverse, les radicaux hydroxyles possèdent une réactivité chimique extrême et oxydent instantanément l'ADN, les protéines et les lipides membranaires environnants dès leur formation.
La simple détection de niveaux élevés de ROS totaux ne suffit pas à elle seule pour mener des recherches rigoureuses sur le stress oxydatif. La surexpression des ROS peut provenir de stimuli cellulaires distincts de durée variable, et les cascades de signalisation en aval présentent des cinétiques d'activation décalées. Pour obtenir des données expérimentales complètes et reproductibles, les chercheurs ont besoin d'une panoplie complète d'outils de recherche comprenant des modèles de stress cellulaire stables, des kits de détection biochimique multi-indices et des réactifs d'analyse spécifiques aux voies de signalisation – autant d'éléments fournis intégralement par Solarbio pour simplifier votre profilage redox multidimensionnel.
Sources de ROS à l'intérieur et à l'extérieur de la cellule
Les mitochondries constituent la principale source endogène d'espèces réactives de l'oxygène (ROS). Lors de la phosphorylation oxydative mitochondriale, les fuites d'électrons des complexes I et III de la chaîne de transport d'électrons permettent aux électrons libres de réagir avec l'oxygène moléculaire pour générer des radicaux superoxydes. Le réticulum endoplasmique produit également des ROS en réponse au stress lié aux protéines mal repliées. La β-oxydation des acides gras peroxysomaux et l'activation de la NADPH oxydase (principale source de ROS dans les modèles cellulaires immunitaires et inflammatoires) contribuent également aux réserves intracellulaires de ROS.
Les facteurs de stress exogènes peuvent augmenter considérablement la production d'espèces réactives de l'oxygène (ROS) cellulaires : les rayonnements ultraviolets et ionisants induisent directement la formation de radicaux ; les métaux lourds inhibent l'activité des enzymes antioxydantes et perturbent la fonction mitochondriale ; les substances toxiques liées au tabac, l'éthanol en excès, les particules fines en suspension dans l'air et les agents chimiothérapeutiques perturbent collectivement l'homéostasie redox. Il est important de noter que les conditions de culture cellulaire de routine peuvent modifier les niveaux basaux de ROS et introduire un biais expérimental, notamment la formulation du milieu de base, le lot de sérum de veau fœtal, le nombre de passages cellulaires et la densité d'ensemencement. Ces variables, souvent négligées, entraînent fréquemment des divergences importantes entre les groupes parallèles. Solarbio fournit des milieux de culture cellulaire standardisés, du sérum à faible teneur en endotoxines et des réactifs de dissociation cellulaire uniformes afin de minimiser ces interférences expérimentales.
Comment les ROS endommagent-elles les cellules ?
Normalement, les ROS fonctionnent comme des molécules de signalisation. Cependant, à partir d'un certain seuil, elles peuvent commencer à endommager des composants cellulaires vitaux.
Dommages à l'ADN
Les radicaux hydroxyles, très réactifs, induisent une modification oxydative des bases guanine de l'ADN, formant ainsi le 8-OHdG, biomarqueur de référence des lésions oxydatives de l'ADN. Ces radicaux provoquent également des cassures simple et double brin de l'ADN, ainsi que des pontages irréversibles entre l'ADN et les protéines. Les lésions transitoires de l'ADN entraînent un arrêt temporaire du cycle cellulaire et activent les voies endogènes de réparation de l'ADN. En revanche, les dommages oxydatifs persistants ou récurrents sont responsables de l'instabilité génomique, de la sénescence cellulaire et de la transformation maligne.
Oxydation des protéines
Les ROS oxydent directement les chaînes latérales des acides aminés au sein des polypeptides : les groupements sulfhydryles de la cystéine subissent une oxydation réversible, tandis que les groupements méthylènes de la méthionine forment des doubles liaisons carbone-carbone après oxydation. L’intensité de l’oxydation produit des effets fonctionnels diamétralement opposés : une légère oxydation de la cystéine agit comme un interrupteur redox réversible pour la transduction du signal, tandis qu’une modification oxydative importante déclenche un repliement protéique incorrect irréversible, une inactivation fonctionnelle et la formation d’agrégats insolubles. Cette voie est centrale dans la recherche sur la neurodégénérescence : une surcharge prolongée en ROS altère les systèmes d’élimination des protéines endommagées et accélère l’agrégation des plaques β-amyloïdes, une caractéristique pathologique majeure de la maladie d’Alzheimer.
Peroxydation lipidique
Les membranes plasmiques et organellaires sont riches en acides gras polyinsaturés, particulièrement sensibles à l'attaque oxydative par les espèces réactives de l'oxygène (ROS). La peroxydation lipidique génère des produits finaux stables, le malondialdéhyde (MDA) et le 4-hydroxynonénal (4-HNE), les biomarqueurs les plus fréquemment quantifiés des lésions oxydatives lipidiques. Ces aldéhydes toxiques perturbent l'intégrité membranaire, altèrent la fonction mitochondriale, se lient aux protéines et à l'ADN cellulaires et créent une boucle de rétroaction positive amplifiant la signalisation inflammatoire. Afin d'obtenir des données expérimentales robustes et multidimensionnelles, la plupart des équipes de recherche mesurent simultanément l'abondance des ROS, les niveaux de peroxydation lipidique, l'activité des enzymes antioxydantes, la sécrétion de cytokines pro-inflammatoires et la viabilité cellulaire.
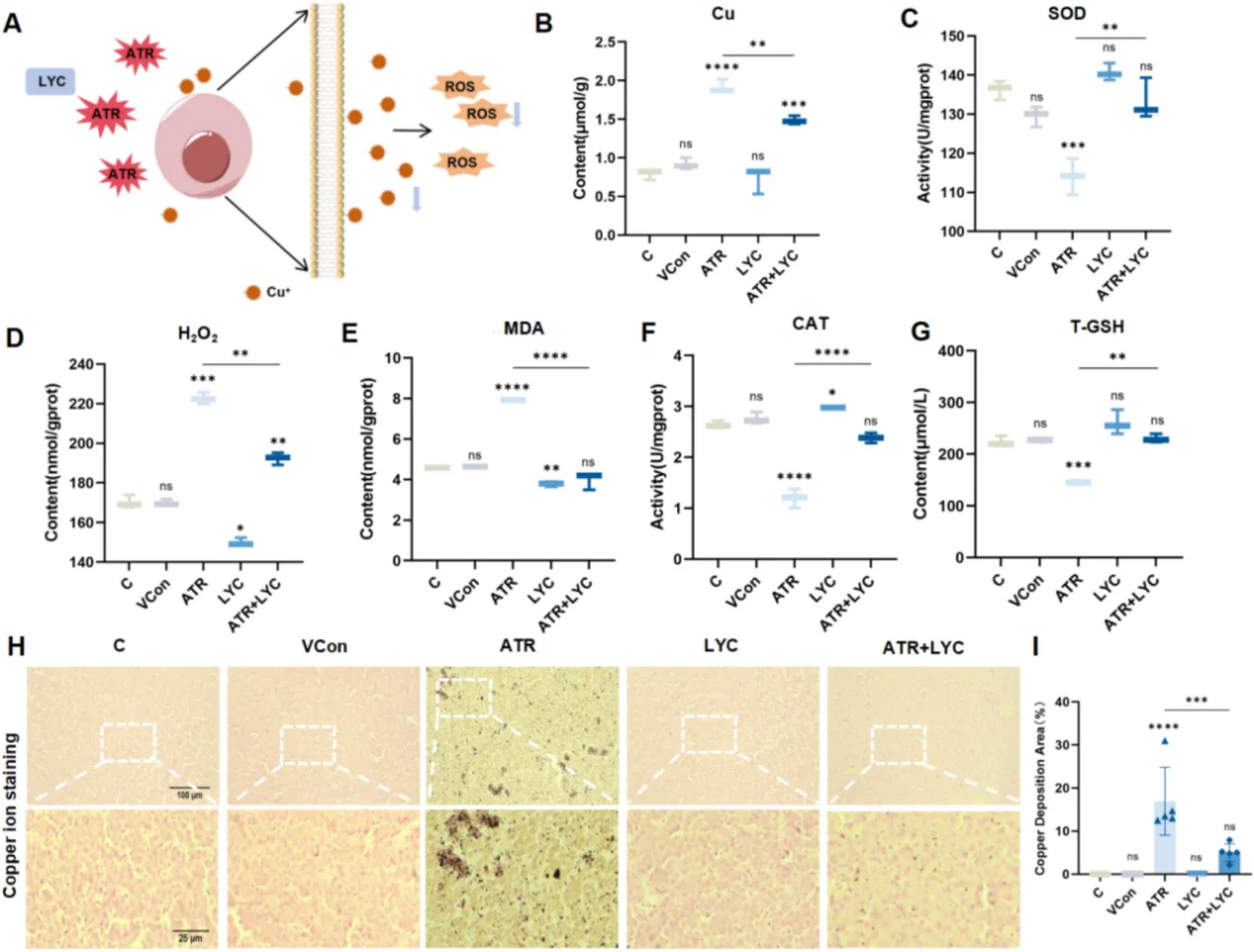
Principales voies de signalisation dans le stress oxydatif
Le stress oxydatif ne se traduit pas uniquement par une seule série d'événements cellulaires ultérieurs. Les ROS peuvent endommager des molécules individuelles et activer ou moduler… voies de transduction du signal, diverses réponses cellulaires différentes peuvent être déclenchées par le stress oxydatif, qui dépendent à leur tour de l'intensité et de la durée du stress oxydatif ainsi que du contexte cellulaire.
Voie Nrf2-ARE : Défense antioxydante
La cascade Nrf2-ARE constitue la principale voie de régulation de la défense antioxydante cellulaire. Dans des conditions physiologiques d'homéostasie redox, la protéine Nrf2 cytoplasmique se lie fortement à Keap1 et subit une ubiquitination et une dégradation protéasomale continues. En cas d'accumulation d'espèces réactives de l'oxygène (ROS), les radicaux libres modifient de manière covalente les résidus cystéine de Keap1, altérant sa conformation et entraînant la dissociation de Nrf2. Simultanément, la voie PI3K/Akt activée phosphoryle Nrf2, accélérant ainsi sa translocation nucléaire. Dans le noyau, Nrf2 forme des hétérodimères avec les petites protéines Maf et se lie aux éléments de réponse antioxydante (ARE) présents sur les promoteurs des gènes cibles.
Ce programme transcriptionnel stimule l'expression des enzymes antioxydantes (SOD, CAT, GSH-Px), des enzymes de synthèse du glutathion et des protéines de détoxification de phase II, renforçant ainsi la capacité d'élimination des ROS et la réparation des dommages cellulaires. De nombreux modèles précliniques confirment que l'activation de Nrf2 exerce des effets cytoprotecteurs dans les lésions hépatiques, pulmonaires, neuronales et épithéliales.
Il est à noter que les résultats des voies de signalisation présentent une dynamique temporelle asynchrone : la translocation nucléaire de Nrf2, l’expression de l’ARNm/protéine du gène cible en aval, l’activité des enzymes antioxydantes et l’efficacité d’élimination des ROS n’atteignent pas simultanément des valeurs maximales.
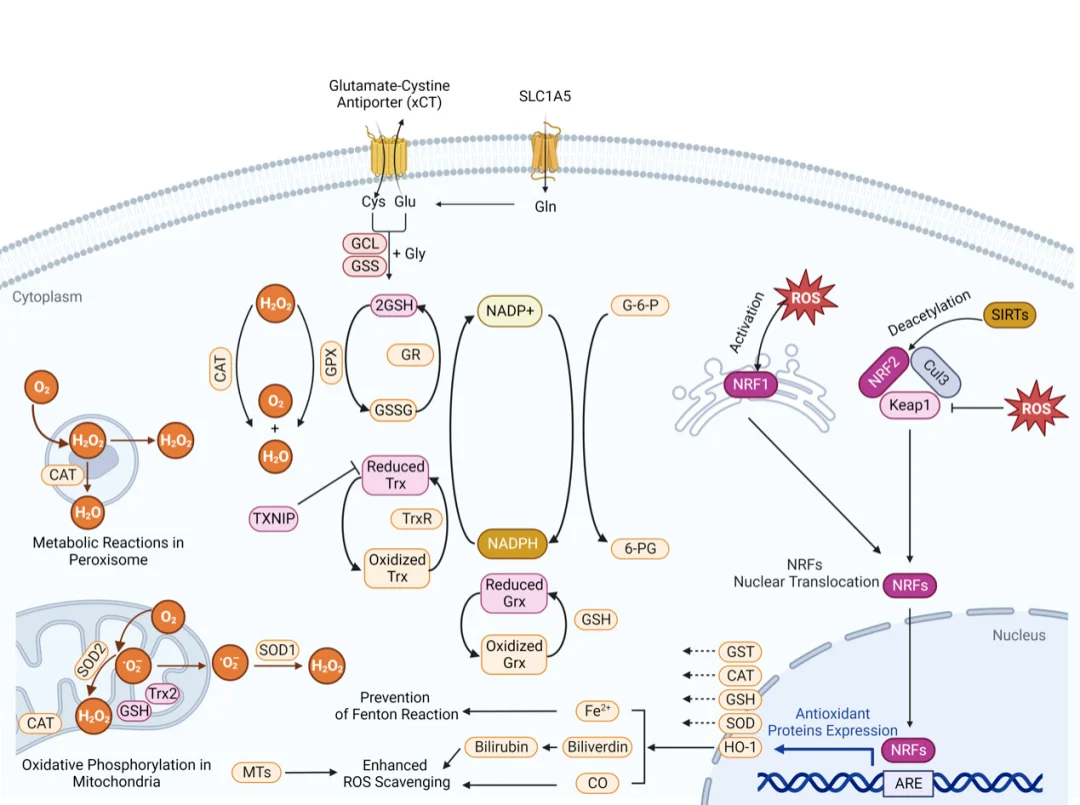
Voie NF-κB : un carrefour d’interactions entre le stress oxydatif et l’inflammation
La cascade NF-κB constitue le lien moléculaire essentiel entre la surcharge en espèces réactives de l'oxygène (ROS) et les réponses inflammatoires. Dans les cellules au repos non stimulées, les dimères NF-κB se lient fortement aux protéines inhibitrices IκB et restent séquestrés dans le cytoplasme. L'augmentation des ROS active le complexe kinase IKK, qui phosphoryle IκB, déclenchant ainsi son ubiquitination et sa dégradation par le protéasome. Le NF-κB libre migre alors vers le noyau et induit la transcription de médiateurs pro-inflammatoires, notamment le TNF-α, l'IL-1β et l'IL-6.
Cette voie forme un cercle vicieux d'auto-amplification : les ROS initient la signalisation inflammatoire, et les cellules immunitaires activées sécrètent des ROS supplémentaires pour stimuler davantage l'activité de NF-κB. Cette activation cyclique persistante favorise la progression des maladies inflammatoires chroniques, notamment l'athérosclérose, la polyarthrite rhumatoïde, les lésions oxydatives hépatiques et l'inflammation métabolique de bas grade.
Cascade MAPK : Régulateur principal de la prolifération cellulaire, de la réponse au stress et de l’apoptose
Les cascades de MAP kinases (MAPK) sont activées par divers stress cellulaires, notamment les ROS (espèces réactives de l'oxygène). Elles comprennent trois sous-familles fonctionnellement distinctes : ERK, JNK et p38 MAPK, qui agissent par phosphorylation séquentielle médiée par MKK. De faibles concentrations physiologiques de ROS activent sélectivement la voie de signalisation ERK, favorisant ainsi la progression du cycle cellulaire, la prolifération et l'homéostasie redox adaptative. À l'inverse, une accumulation excessive de ROS induit une forte phosphorylation de JNK et de p38, déclenchant des réponses au stress cellulaire, la transcription de gènes pro-inflammatoires et la mort cellulaire par apoptose.
Dans les modèles de lésions d'ischémie-reperfusion myocardique, la réintroduction rapide d'oxygène induit une forte production d'espèces réactives de l'oxygène (ROS), hyperactivant la voie de signalisation JNK/p38 et déclenchant une apoptose massive des cardiomyocytes. En oncologie, de faibles concentrations basales de ROS favorisent la prolifération tumorale via l'activation d'ERK, tandis que l'augmentation pharmacologique des niveaux de ROS sensibilise les cellules malignes à la chimio- et à la radiothérapie. Ainsi, la concentration de ROS et la durée d'exposition déterminent le devenir cellulaire final régulé par la voie de signalisation MAPK.
Voie PI3K/Akt : Signalisation de survie cytoprotectrice sous stress redox modéré
La cascade PI3K/Akt constitue le principal axe de signalisation de survie cellulaire en cas de stress oxydatif modéré. Une stimulation modérée par les ROS active la PI3K, générant du PIP3 membranaire qui recrute l'Akt cytoplasmique à la membrane plasmique pour sa phosphorylation et son activation complète. L'Akt phosphorylée inhibe la signalisation apoptotique par de multiples mécanismes : elle phosphoryle et inactive la protéine pro-apoptotique Bad, inhibe l'activation de la cascade des caspases et stimule le métabolisme et la prolifération cellulaires via mTOR. De plus, l'Akt phosphoryle directement Nrf2, accélérant ainsi sa translocation nucléaire et amplifiant les réponses transcriptionnelles antioxydantes.
Cette signalisation protectrice présente un seuil redox critique. Lorsque les concentrations d'espèces réactives de l'oxygène (ROS) dépassent le seuil tolérable, les protéines PI3K et Akt subissent une inactivation oxydative irréversible et une dégradation protéolytique, ce qui oriente la signalisation cellulaire vers la voie apoptotique intrinsèque mitochondriale. Dans les modèles neuronaux de la maladie de Parkinson, l'efficacité thérapeutique des interventions antioxydantes dépend fortement de l'intégrité fonctionnelle de la voie de signalisation PI3K/Akt.
Liens entre maladies et recherche sur le stress oxydatif
Le stress oxydatif constitue un facteur pathologique commun à de nombreuses maladies humaines. Plutôt que de se contenter d'identifier les phénotypes de lésions cellulaires, l'étude approfondie des cascades de signalisation redox sous-jacentes qui déclenchent les lésions tissulaires présente une valeur translationnelle bien plus importante.
Recherche sur le cancer
Les cellules tumorales présentent un statut ROS basal hétérogène : certains sous-types maintiennent des niveaux de ROS intrinsèquement élevés, tandis que d’autres activent l’axe antioxydant Nrf2 pour supprimer l’accumulation de ROS, ce qui leur confère une forte résistance à la chimio- et à la radiothérapie. Cependant, l’augmentation artificielle des ROS intracellulaires jusqu’à des seuils toxiques peut induire la mort des cellules malignes. En recherche oncologique, les systèmes de détection multidimensionnels sont essentiels et comprennent des sondes fluorescentes pour les ROS, des kits de tests de sensibilité aux médicaments, des chimiothèques de petites molécules et des anticorps biomarqueurs spécifiques de voies de signalisation.
Recherche sur les maladies neurodégénératives
Les neurones matures sont des cellules post-mitotiques incapables de se régénérer, caractérisées par un métabolisme énergétique fragile et une extrême vulnérabilité au stress oxydatif. L'altération des mitochondries et des systèmes de repliement des protéines induite par les espèces réactives de l'oxygène (ROS) déclenche l'apoptose, une mort cellulaire programmée. Des modèles animaux de la maladie de Parkinson montrent qu'un excès de ROS supprime simultanément la voie cytoprotectrice PI3K/Akt et hyperactive la voie pro-apoptotique JNK. Dans la maladie d'Alzheimer, la surcharge en ROS accélère l'agrégation pathologique du peptide β-amyloïde et induit une neuroinflammation persistante ; l'activation ciblée de Nrf2 peut atténuer efficacement ces phénotypes neurodégénératifs.
Modèles de lésions cardiovasculaires et hépatiques
En recherche cardiovasculaire, l'activation persistante de NF-κB induit une inflammation endothéliale et une surexpression des molécules d'adhésion, accélérant la formation de plaques d'athérosclérose. Une forte production d'espèces réactives de l'oxygène (ROS) consécutive à une lésion d'ischémie-reperfusion myocardique active les cascades JNK/p38 MAPK et déclenche une apoptose massive des cardiomyocytes. Dans les modèles de toxicité hépatique, le stress oxydatif induit un dysfonctionnement mitochondrial, une peroxydation lipidique importante, la sécrétion de cytokines pro-inflammatoires et une cuproptose. Un protocole de recherche classique utilise l'atrazine pour induire des lésions oxydatives hépatiques, le lycopène agissant comme agent antioxydant pour réparer les tissus endommagés.
Instructions d'utilisation du produit pour les expériences sur le stress oxydatif
Pour les expériences de sauvetage antioxydant, nous recommandons la N-acétyl-L-cystéine (NAC) (réf. IA0050), un modulateur redox classique à petite molécule utilisé dans la recherche sur les ROS. La NAC reconstitue les réserves intracellulaires de thiols et stimule les systèmes antioxydants dépendants du glutathion, permettant ainsi aux chercheurs de vérifier si les changements phénotypiques observés sont dus à un stress induit par les ROS ou à des effets hors cible non pertinents.
Le lycopène (réf. IL0510) constitue un autre agent antioxydant fiable pour les essais cellulaires in vitro et les modèles de tissus animaux in vivo, notamment pour la recherche sur les lésions hépatiques et la peroxydation lipidique. Une évaluation complète de l'efficacité antioxydante nécessite la détection de plusieurs paramètres à différentes concentrations, en tenant compte des interférences du solvant, des niveaux d'espèces réactives de l'oxygène (ROS), de l'activité antioxydante endogène et de la viabilité cellulaire ; négliger l'un de ces paramètres conduirait à des conclusions mécanistiques incomplètes et unilatérales.
Avant de lancer un test de détection du stress oxydatif, les chercheurs doivent confirmer les conditions de stockage des réactifs, la solubilité du solvant, la certification de la qualité des lots et les fenêtres temporelles de détection valides. Les résultats expérimentaux sont facilement faussés par la photodégradation, les cycles de congélation-décongélation répétés et les protocoles de lyse cellulaire non uniformes. Des informations sur la mise en place d'un test ou les produits compatibles sont disponibles sur le site web. page Web des services techniques de Solarbio ou en parcourant les données pertinentes articles techniques.
Conclusion
Le stress oxydatif ne se résume pas à une simple accumulation excessive d'espèces réactives de l'oxygène (ROS). Il résulte d'un réseau d'interactions complexe : une production non régulée de ROS sature les systèmes d'élimination antioxydants endogènes, entraînant des modifications irréversibles de l'ADN, des protéines et des lipides membranaires. Les métabolites lipidiques oxydés amplifient les cascades inflammatoires, qui à leur tour aggravent les dommages oxydatifs et induisent finalement la mort cellulaire par apoptose.
Les cellules déploient quatre axes de signalisation principaux pour répondre au déséquilibre redox : la voie Nrf2-ARE initie la défense antioxydante globale ; NF-κB intervient dans les réponses inflammatoires déclenchées par le stress oxydatif ; les cascades MAPK déterminent la prolifération ou l’apoptose cellulaire en fonction du dosage de ROS ; la voie PI3K/Akt maintient la survie cellulaire en cas de perturbation redox légère.
La détection d'un seul indicateur ne permet pas de mener des recherches rigoureuses sur le stress oxydatif. Pour obtenir des données complètes et reproductibles, la détection simultanée de plusieurs indicateurs est fortement recommandée, notamment la concentration totale de ROS, l'activité des enzymes antioxydantes, les niveaux de peroxydation lipidique MDA/4-HNE, l'état de phosphorylation des principales protéines de signalisation, la sécrétion de cytokines pro-inflammatoires, la fonction mitochondriale, les biomarqueurs liés à l'apoptose et la viabilité cellulaire. Si vous avez besoin d'un produit adapté à vos besoins, de protocoles expérimentaux détaillés ou d'une assistance technique personnalisée, n'hésitez pas à nous contacter. L'équipe de recherche professionnelle de Solarbio.
FAQ (questions fréquentes)
Q1 : Qu'est-ce que le stress oxydatif exactement ?
A1 : Le stress oxydatif désigne une perturbation de l’homéostasie redox intracellulaire, déclenchée par une production excessive d’espèces réactives de l’oxygène (ROS) ou une capacité d’élimination des antioxydants altérée. Ce déséquilibre induit des dommages biomoléculaires, une transduction aberrante du signal, des réponses inflammatoires et la mort cellulaire par apoptose, ou une combinaison de ces phénotypes pathologiques.
Q2 : Les ROS sont-elles intrinsèquement nocives pour les cellules ?
A2 : Non. Les ROS, à faible concentration et de façon transitoire, agissent comme messagers physiologiques essentiels. Seuls des niveaux élevés et soutenus de ROS, ou une surcharge localisée de ROS au sein d'organites sensibles à l'oxydoréduction (mitochondries, noyau), exerceront des effets cytotoxiques.
Q3 : Quelle voie de signalisation centrale domine la défense antioxydante cellulaire ?
A3 : La voie Nrf2-ARE constitue l’axe de régulation antioxydant principal. Elle stimule la transcription de la SOD, de la CAT, de la GSH-Px, des enzymes de synthèse dépendantes du glutathion et des protéines de détoxification de phase II afin de rétablir l’équilibre redox.
Q4 : Pourquoi NF-κB est-il une cible de recherche essentielle pour les études sur le stress oxydatif ?
A4 : NF-κB est le principal lien moléculaire entre la surcharge en ROS et l’inflammation. Il forme une boucle de rétroaction positive auto-amplificatrice : les ROS activent NF-κB pour induire l’inflammation, et les cellules immunitaires activées sécrètent davantage de ROS pour maintenir l’activation de la voie, aggravant ainsi les lésions tissulaires chroniques.
Q5 : Quelles sont les principales précautions à prendre lors de l’application de NAC dans les expériences sur les ROS ?
A5 : Les chercheurs doivent optimiser au préalable la concentration de NAC, la durée d’incubation et la formulation du solvant, et réaliser des tests de viabilité cellulaire en parallèle afin d’exclure toute interférence cytotoxique. La co-détection multi-indices de biomarqueurs redox est nécessaire pour obtenir des conclusions expérimentales solides et crédibles.



