






酸化ストレスの分子メカニズムとシグナル伝達経路:細胞内で実際に何が起きているのか
目次
酸化ストレスは、細胞生物学、毒性学、薬理学、老年科学、腫瘍学、神経変性疾患モデリングなど、幅広い分野で研究の中心的なテーマとなっています。その基本的な定義は単純明快に聞こえます。細胞内の活性酸素種(ROS)が細胞本来の抗酸化物質除去能力を超えるレベルまで蓄積し、酸化還元恒常性を阻害する、というものです。しかし、実際の細胞培養実験では、はるかに複雑な制御ロジックが明らかになっています。軽度で一時的なROSバーストは、生理学的シグナル伝達を媒介する重要なセカンドメッセンジャーとして機能しますが、持続的な高濃度のROS過負荷は、DNA、タンパク質、膜脂質に不可逆的な損傷を引き起こし、ミトコンドリア機能障害、炎症カスケード、アポトーシス細胞死、多様な細胞ストレス応答を順次引き起こします。
ROS蛍光検出、抗酸化効果スクリーニング、炎症性ストレス経路研究を行う研究室にとって、ROS上昇の程度、持続時間、細胞内局在を定量化することは、単に一般的なROS上昇を確認するよりもはるかに重要です。細胞株の由来、薬剤のインキュベーション時間、蛍光プローブの負荷効率、試薬のグレード、コールドチェーン保管プロトコル、処理後のサンプル処理など、複数の交絡因子が実験結果を大きく歪める可能性があります。 ソレボ 生化学試薬、低分子生物活性化合物、一次/二次抗体、ELISA検出キット、細胞培養消耗品など、ライフサイエンス研究に必要なあらゆる資材を提供し、分子生物学、免疫学、生化学経路の研究ニーズに完全に対応し、研究者が実験におけるバイアスを最小限に抑えられるよう支援します。
酸化ストレスとは何か?
酸化ストレスとは、過剰な活性酸素種(ROS)の生成、あるいは抗酸化物質の解毒能力の低下によって引き起こされる、細胞内における酸化還元平衡状態の乱れを指します。持続的に高濃度のROSが存在すると深刻な細胞毒性を示す一方、低濃度で一時的なROSプールは、免疫活性化、細胞増殖、組織発生過程を調節する重要なシグナル伝達メディエーターとして機能します。酸化ストレスの発生は、ROSの過剰な蓄積、酸化還元感受性細胞内区画(ミトコンドリアと核)におけるROSの長期にわたる過負荷、そして長期にわたる酸化還元状態の不均衡という3つの主要な条件によって引き起こされ、これらすべてが下流の細胞毒性カスケードを誘発します。
主なROSの種類
真核細胞内には多様な活性酸素種が存在する。最も広く研究されている3つのサブタイプはスーパーオキシドアニオンラジカル(O2–•)、過酸化水素(H2O2)、およびヒドロキシルラジカル(•おお)スーパーオキシドアニオンラジカルは、主にミトコンドリアの電子伝達系で生成される。 H2O2 は優れた化学的安定性と高い膜透過性を示し、細胞小器官の境界を越えて移動し、酸化還元シグナル伝達物質および細胞ストレスセンサーの両方として機能する。これとは対照的に、ヒドロキシルラジカルは極めて高い化学反応性を持ち、生成されるとすぐに周囲のDNA、タンパク質、膜脂質を酸化する。
単にROS総量の上昇を検出するだけでは、厳密な酸化ストレス研究を支えることはできません。ROSの上昇は、持続時間の異なる様々な細胞刺激に起因する可能性があり、下流のシグナル伝達カスケードは活性化のタイミングがずれています。包括的で再現性のある実験データを取得するには、研究者は安定した細胞ストレスモデル、マルチインデックス生化学検出キット、および経路特異的分析試薬を網羅した完全な研究ツールキットを必要とします。これらはすべて、当社によって完全に供給されています。 ソレボ 多次元酸化還元プロファイリングを簡素化するため。
細胞内外の活性酸素種発生源
ミトコンドリアは、主要な内因性活性酸素種(ROS)発生源である。ミトコンドリアの酸化的リン酸化の過程で、電子伝達系複合体Iおよび複合体IIIからの電子漏出により、遊離電子が分子状酸素と反応してスーパーオキシドラジカルを生成する。小胞体もまた、タンパク質折り畳み異常応答ストレス下でROSを生成する。ペルオキシソームにおける脂肪酸β酸化およびNADPHオキシダーゼの活性化(免疫細胞および炎症細胞モデルにおける主要なROS発生源)も、細胞内ROSプールの増加に寄与する。
外因性ストレス因子は細胞内ROS量を劇的に増加させる可能性がある。紫外線や電離放射線はラジカル生成を直接誘導し、重金属は抗酸化酵素活性を抑制しミトコンドリア機能を阻害する。タバコ関連毒性物質、過剰なエタノール、大気浮遊粒子状物質、化学療法剤は、酸化還元恒常性を総合的に破壊する。特に、通常の細胞培養条件は、基礎培地組成、ウシ胎児血清のロット、細胞継代数、播種密度などを含め、基礎ROSレベルを変化させ、実験バイアスをもたらす可能性がある。これらの見落とされがちな変数は、並行群間で大きな差異を生じさせることが多い。 ソレボ 標準化された細胞培養培地、低エンドトキシン血清、および均一な細胞解離試薬を提供することで、このような実験上の干渉を最小限に抑えます。
ROSはどのように細胞にダメージを与えるのか?
通常、活性酸素種(ROS)はシグナル分子として機能する。しかし、ある一定量を超えると、細胞の重要な構成要素に損傷を与え始める可能性がある。
DNA損傷
高反応性のヒドロキシルラジカルは、DNAグアニン塩基の酸化修飾を引き起こし、酸化DNA損傷の典型的なバイオマーカーである8-OHdGを生成する。これらのラジカルはまた、一本鎖および二本鎖DNA切断、ならびに不可逆的なDNA-タンパク質架橋も誘発する。一過性のDNA損傷は一時的な細胞周期停止を引き起こし、内因性DNA修復経路を活性化するが、持続的または反復的な酸化損傷はゲノム不安定性、細胞老化、および悪性形質転換を促進する。
タンパク質酸化
ROSはポリペプチド内のアミノ酸側鎖を直接酸化します。システインのスルフィドリル基は可逆的に酸化されますが、メチオニンのメチレン基は酸化後に炭素-炭素二重結合を形成します。酸化の強度は正反対の機能的結果をもたらします。軽度のシステイン酸化はシグナル伝達の可逆的な酸化還元スイッチとして機能しますが、強力な酸化修飾は不可逆的なタンパク質ミスフォールディング、機能不活性化、および不溶性凝集体の形成を引き起こします。この経路は神経変性研究の中心であり、持続的なROS過負荷は損傷タンパク質のクリアランスシステムを損ない、アルツハイマー病の特徴的な病理学的特徴であるβアミロイドプラークの凝集を促進します。
脂質過酸化
血漿膜および細胞小器官膜は多価不飽和脂肪酸に富んでおり、これらは活性酸素種(ROS)による酸化攻撃を受けやすい。脂質過酸化により、安定な最終生成物であるマロンジアルデヒド(MDA)と4-ヒドロキシノネナール(4-HNE)が生成される。これらは脂質酸化損傷の最も一般的な定量バイオマーカーである。これらの毒性アルデヒドは膜の完全性を破壊し、ミトコンドリア機能を損ない、細胞タンパク質やDNAと架橋結合し、炎症シグナルを増幅する正のフィードバックループを形成する。堅牢で多次元的な実験データを得るために、ほとんどの研究グループは、ROS量、脂質過酸化レベル、抗酸化酵素活性、炎症性サイトカイン分泌、細胞生存率を同時に検出している。
酸化ストレスにおける主要なシグナル伝達経路
酸化ストレスは、一連の細胞イベントにそのまま結びつくわけではありません。ROSは個々の分子に損傷を与えるだけでなく、活性化や調節も引き起こす可能性があります。 シグナル伝達経路酸化ストレスによって様々な細胞応答が引き起こされる可能性があり、それらの応答は酸化ストレスの強度と持続時間、そして細胞の背景によって左右される。
Nrf2-ARE経路:抗酸化防御
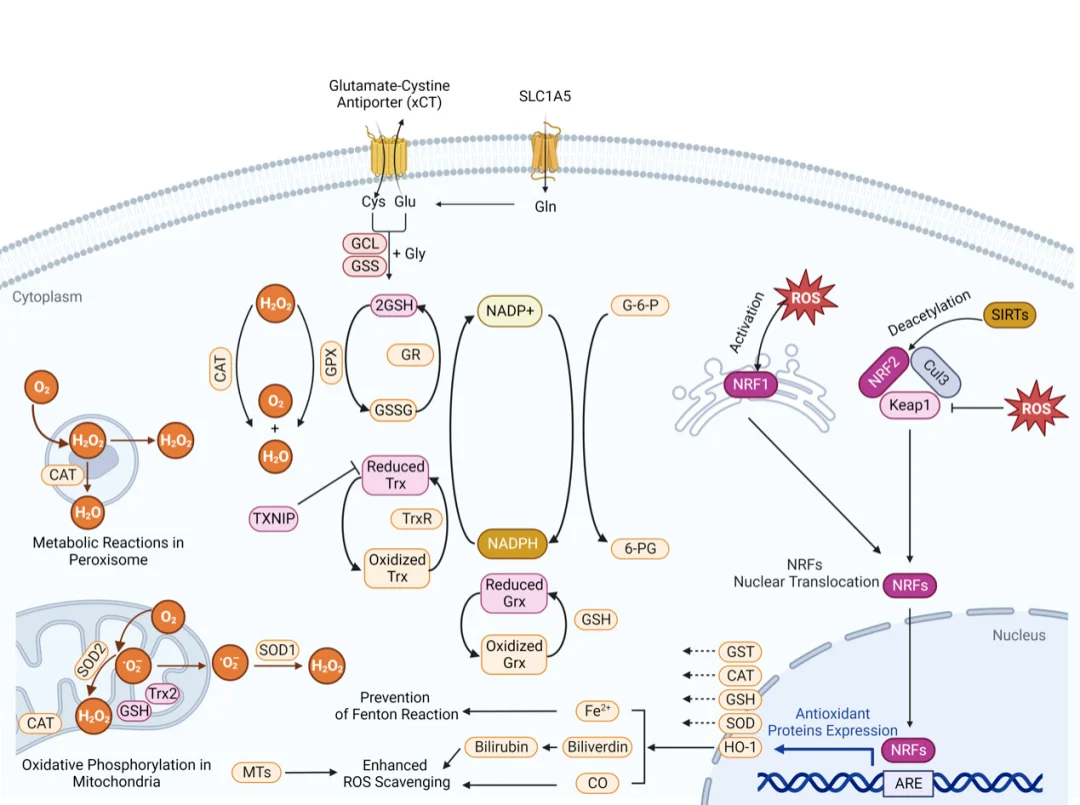
Nrf2-AREカスケードは、細胞の抗酸化防御を制御する主要な調節経路として機能します。生理的な酸化還元恒常性下では、細胞質Nrf2はKeap1に強く結合し、継続的なユビキチン化とプロテアソーム分解を受けます。ROSが蓄積すると、フリーラジカルがKeap1のシステイン残基を共有結合的に修飾し、その立体構造を変化させてNrf2を解離させます。同時に、活性化されたPI3K/AktがNrf2をリン酸化して、核への移行をさらに促進します。核内では、Nrf2は小型Mafタンパク質とヘテロ二量体を形成し、標的遺伝子プロモーター上の抗酸化応答エレメント(ARE)に結合します。
この転写プログラムは、抗酸化酵素(SOD、CAT、GSH-Px)、グルタチオン合成酵素、および第II相解毒タンパク質の発現を促進し、ROS除去能力と細胞損傷修復能力を総合的に向上させます。広範な前臨床モデルにおいて、Nrf2の活性化が肝臓、肺、神経、および上皮組織の損傷に対して細胞保護効果を発揮することが確認されています。
特筆すべきは、経路の読み取り値が非同期的な時間的ダイナミクスを示すことである。Nrf2の核移行、下流の標的遺伝子のmRNA/タンパク質発現、抗酸化酵素活性、およびROS除去効率は、同時にピーク値に達しない。
NF-κB経路:酸化ストレスと炎症のクロストークハブ
NF-κBカスケードは、活性酸素種(ROS)の過剰負荷と炎症反応を結びつける重要な分子的な架け橋として機能します。刺激を受けていない静止細胞では、NF-κB二量体は阻害性タンパク質であるIκBに強く結合し、細胞質内に隔離されたままです。ROSの増加はIKKキナーゼ複合体を活性化し、IκBをリン酸化してユビキチン化とプロテアソーム分解を引き起こします。遊離したNF-κBは核に移行し、TNF-α、IL-1β、IL-6などの炎症性メディエーターの転写を促進します。
この経路は自己増幅的な正のフィードバックの悪循環を形成する。すなわち、活性酸素種(ROS)が炎症シグナル伝達を開始し、活性化された免疫細胞がさらにROSを分泌してNF-κB活性を増強する。この持続的な循環的活性化は、動脈硬化、関節リウマチ、肝臓の酸化損傷、代謝性低度炎症などの慢性炎症性疾患の進行を促進する。
MAPKカスケード:細胞増殖、ストレス応答、アポトーシスのマスターレギュレーター
マイトジェン活性化プロテインキナーゼ(MAPK)カスケードは、活性酸素種(ROS)を含む様々な細胞ストレス因子によって活性化され、ERK、JNK、p38 MAPKという機能的に異なる3つのサブファミリーから構成され、MKKを介した連続的なリン酸化によって機能します。生理的な低濃度のROSは、選択的にERKシグナル伝達を活性化し、細胞周期の進行、増殖、および適応的な酸化還元恒常性を促進します。対照的に、過剰なROS蓄積は、強力なJNKおよびp38リン酸化を引き起こし、細胞ストレス応答、炎症誘発性遺伝子転写、およびアポトーシスによる細胞死を誘発します。
心筋虚血再灌流障害モデルでは、急速な酸素再導入によりROSが急激に増加し、JNK/p38シグナル伝達が過剰に活性化されて心筋細胞のアポトーシスが大量に引き起こされる。腫瘍学研究では、基礎的な低レベルのROSはERK活性化を介して腫瘍の増殖を維持する一方、薬理学的にROSレベルを上昇させると悪性細胞が化学療法や放射線療法に対して感受性が高まる。総合的に見ると、ROS濃度と曝露時間の両方が、MAPKシグナル伝達によって制御される最終的な細胞運命を決定する。
PI3K/Akt経路:中程度の酸化還元ストレス下における細胞保護的な生存シグナル伝達
PI3K/Aktカスケードは、軽度の酸化ストレス下における主要な生存促進シグナル伝達経路である。中程度のROS刺激はPI3Kを活性化し、膜結合型PIP3を生成する。PIP3は細胞質Aktを細胞膜にリクルートし、リン酸化と完全な活性化を促す。リン酸化されたAktは、複数のメカニズムを介してアポトーシスシグナル伝達を抑制する。すなわち、アポトーシス促進タンパク質Badをリン酸化して不活性化し、カスパーゼカスケードの活性化を阻害し、mTORを介した細胞代謝と増殖を促進する。さらに、AktはNrf2を直接リン酸化して核移行を促進し、抗酸化転写応答を増幅する。
この保護シグナル伝達には、重要な酸化還元閾値が存在する。ROS濃度が許容範囲を超えると、PI3KとAktは不可逆的な酸化的不活性化とタンパク質分解を受け、細胞シグナル伝達の優位性がミトコンドリア内在性アポトーシス経路へと移行する。パーキンソン病の神経細胞モデルにおいて、抗酸化介入の治療効果は、PI3K/Aktシグナル伝達の機能状態が維持されているかどうかに大きく依存する。
酸化ストレス研究における疾患との関連性
酸化ストレスは、数多くのヒト疾患に共通する病理学的要因として作用する。細胞損傷の表現型を特定するだけでなく、組織損傷を引き起こす根本的な酸化還元シグナル伝達経路を解明することで、はるかに大きな臨床応用研究上の価値が得られる。
がん研究
腫瘍細胞は、ROSの基礎状態が不均一である。一部のサブタイプは本質的に高いROSレベルを維持する一方、他のサブタイプはNrf2抗酸化経路を活性化してROSの蓄積を抑制し、化学療法や放射線療法に対する強い耐性を獲得する。とはいえ、細胞内ROSを人為的に毒性閾値まで高めることで、悪性細胞の死を誘導できる。腫瘍学研究においては、ROS蛍光プローブ、薬剤感受性試験キット、低分子化合物ライブラリー、経路特異的バイオマーカー抗体など、多次元的な検出システムが不可欠である。
神経変性疾患研究
成熟したニューロンは、再生能力のない分裂後細胞であり、脆弱なエネルギー代謝と酸化ストレスに対する極めて高い脆弱性を特徴としています。ROSを介したミトコンドリアおよびタンパク質フォールディングシステムの障害は、最終的にアポトーシスによるプログラム細胞死を引き起こします。パーキンソン病の動物モデルでは、過剰なROSが細胞保護的なPI3K/Akt経路を抑制し、アポトーシス促進性のJNKシグナル伝達を過剰に活性化することが示されています。アルツハイマー病では、ROSの過剰負荷が病理的なβアミロイド凝集を加速させ、持続的な神経炎症を引き起こします。Nrf2を標的とした活性化は、これらの神経変性表現型を効果的に軽減することができます。
心血管系および肝臓損傷モデル
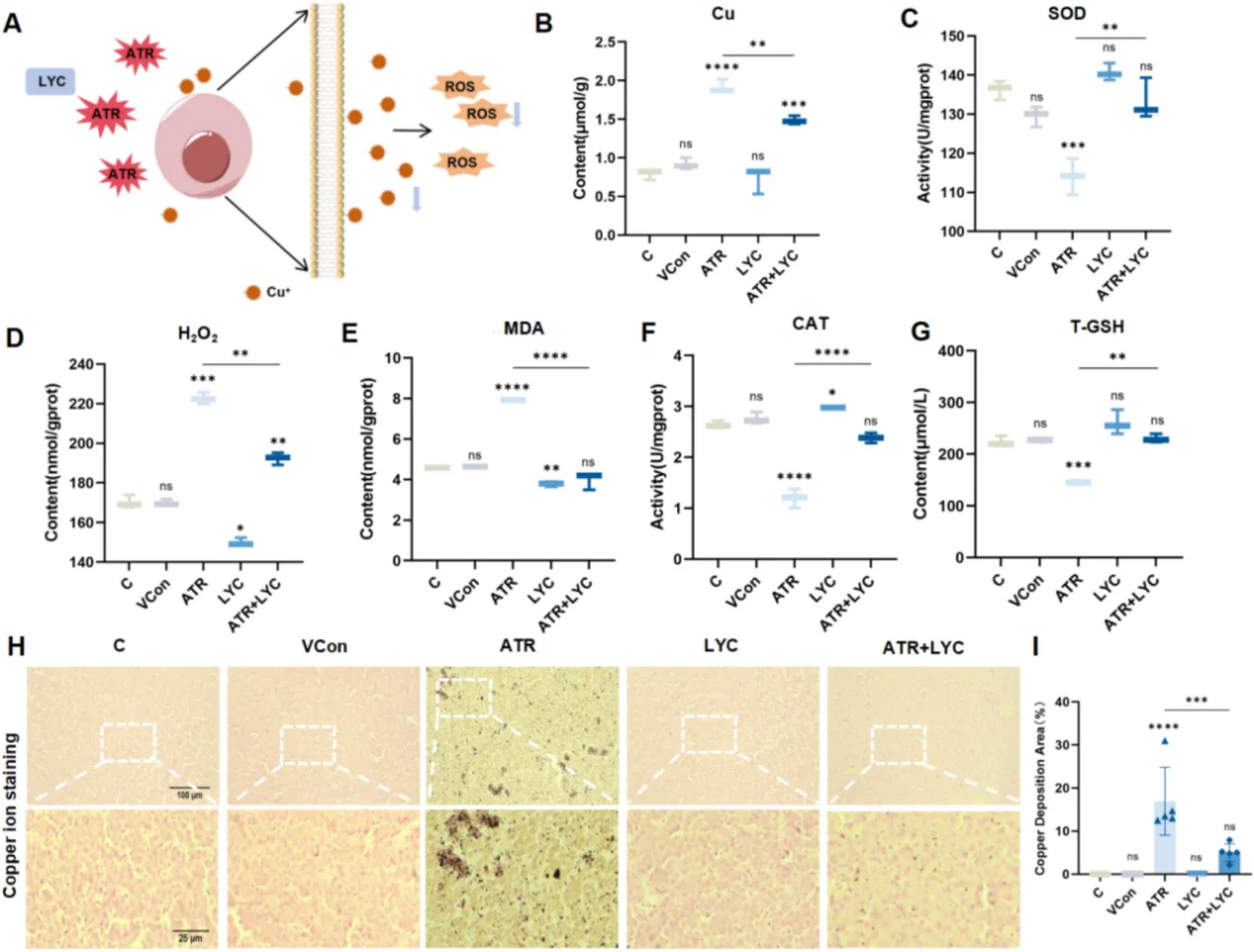
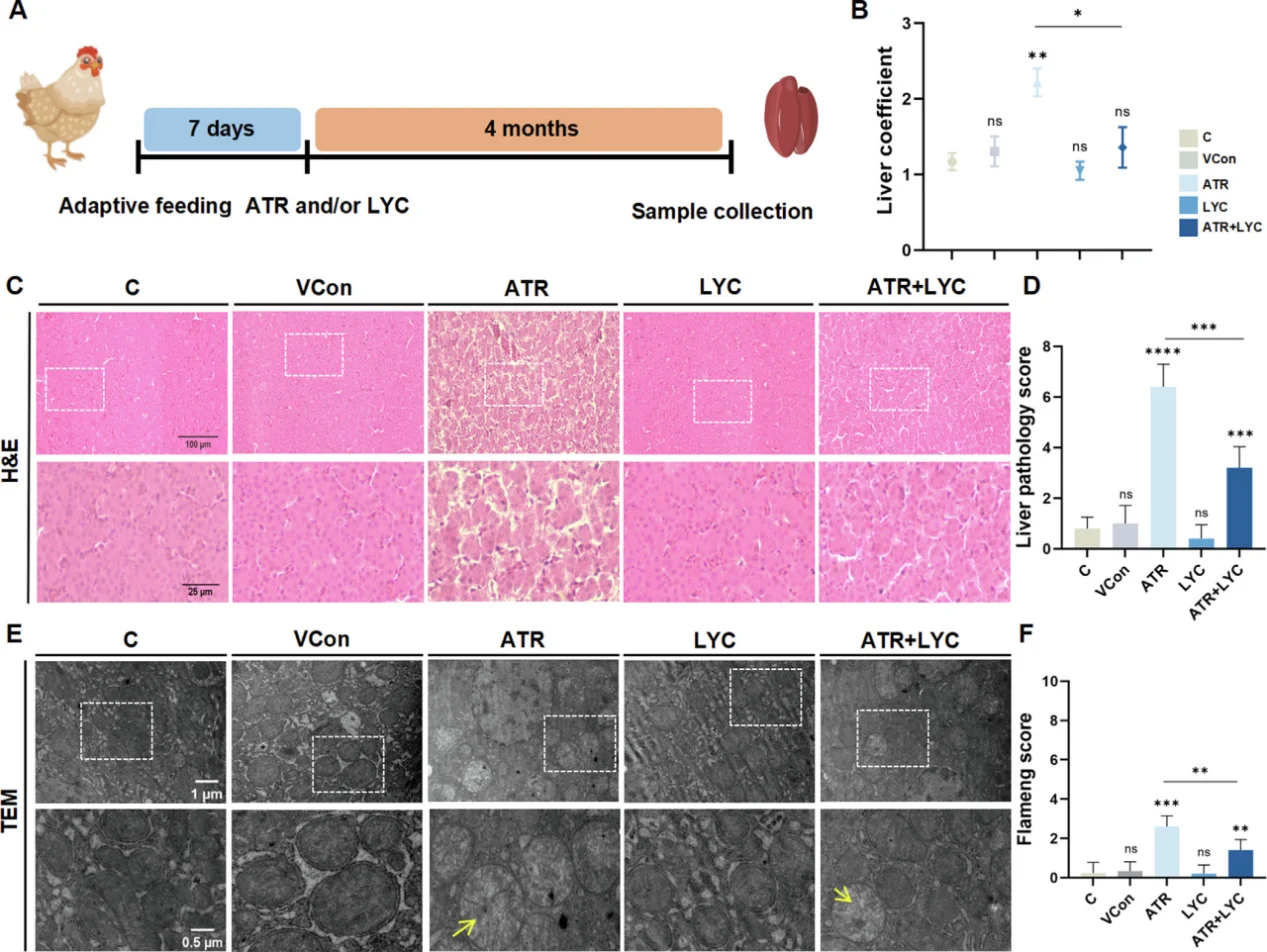
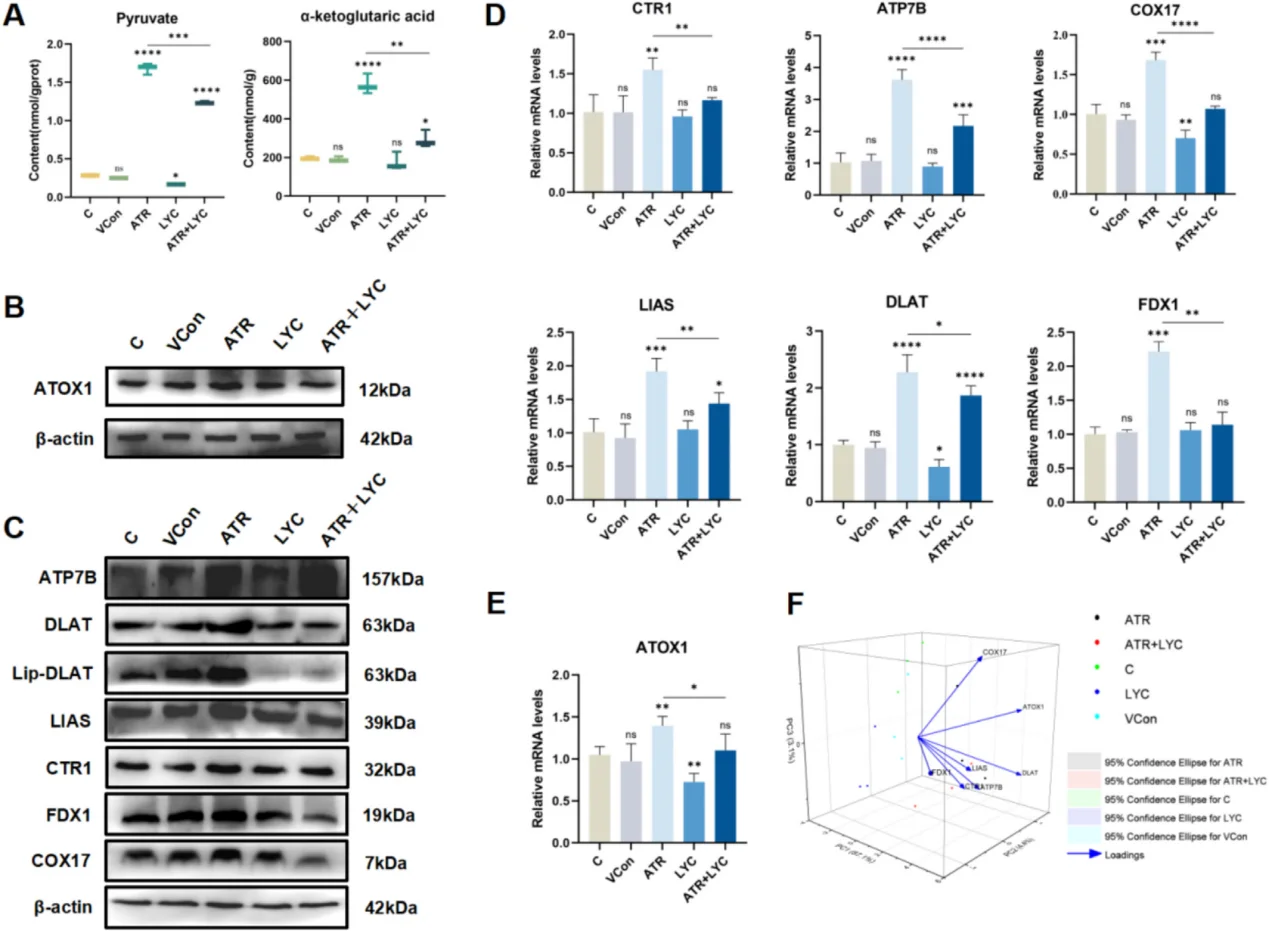
心血管研究において、持続的なNF-κB活性化は内皮細胞の炎症と接着分子の過剰発現を引き起こし、動脈硬化性プラークの形成を加速させる。心筋虚血再灌流障害から生じる急激なROSバーストはJNK/p38 MAPKカスケードを活性化し、心筋細胞の大規模なアポトーシスを引き起こす。肝毒性モデルでは、酸化ストレスはミトコンドリア機能障害、大規模な脂質過酸化、炎症性サイトカイン分泌、およびクプロトーシスを引き起こす。典型的な研究パラダイムでは、アトラジンを用いて肝臓の酸化損傷を誘導し、リコピンを抗酸化介入剤として用いて組織損傷を修復する。
酸化ストレス実験に関する製品説明書
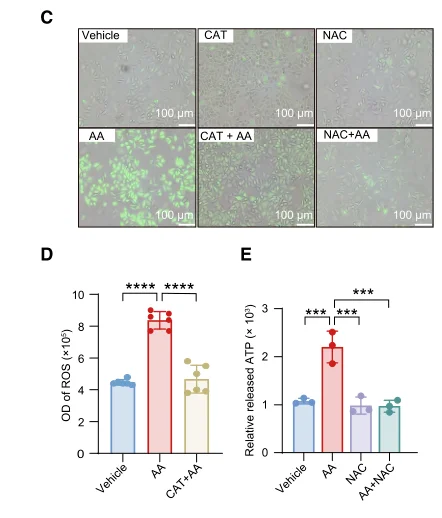
抗酸化物質による救済実験には、ROS研究における定番の低分子レドックス調節剤であるN-アセチル-L-システイン(カタログ番号IA0050)をお勧めします。NACは細胞内チオールプールを補充し、グルタチオン依存性抗酸化システムを強化することで、観察された表現型変化がROSを介したストレスによるものか、あるいは無関係なオフターゲット効果によるものかを確認することを可能にします。
リコピン(カタログ番号IL0510)は、特に肝障害や脂質過酸化の研究において、in vitro細胞アッセイやin vivo動物組織モデルにおける信頼性の高い抗酸化剤として用いられます。抗酸化効果を包括的に評価するには、濃度勾配、溶媒干渉、ROSレベル、内因性抗酸化活性、細胞生存率など、複数の指標を総合的に検出する必要があります。これらの指標のいずれかを無視すると、不完全かつ一方的なメカニズム的結論に至ります。
酸化ストレス検出アッセイを開始する前に、研究者は試薬の保管条件、溶媒の溶解性、バッチ品質認証、および有効な検出時間枠を確認する必要があります。実験結果は、光分解、凍結融解の繰り返し、および一貫性のない細胞溶解プロトコルによって容易に歪められます。アッセイの設定または製品のマッチングに関する情報は、 Solarbioの技術サービスウェブページ または関連商品を閲覧する 技術記事.
結論
酸化ストレスは、単に過剰な活性酸素種(ROS)の蓄積として定義できるものではありません。それは複雑な相互作用ネットワークから生じます。制御されないROS産生は、内因性の抗酸化物質除去システムを圧倒し、DNA、タンパク質、膜脂質の不可逆的な変化を引き起こします。酸化された脂質代謝産物は炎症カスケードをさらに増幅させ、それが酸化損傷を悪化させ、最終的にはアポトーシスによる細胞死を誘導します。
細胞は酸化還元バランスの不均衡に対応するために4つの主要なシグナル伝達軸を展開します。Nrf2-ARE経路は全体的な抗酸化防御を開始し、NF-κBは酸化ストレスによって引き起こされる炎症反応を媒介し、MAPKカスケードはROSの量に基づいて細胞増殖またはアポトーシスを決定し、PI3K/Akt経路は軽度の酸化還元撹乱下での細胞生存を維持します。
単一指標の検出では、厳密な酸化ストレス研究をサポートできません。包括的で再現性のあるデータを取得するには、総ROS濃度、抗酸化酵素活性、MDA/4-HNE脂質過酸化レベル、主要シグナル伝達タンパク質のリン酸化状態、炎症性サイトカイン分泌、ミトコンドリア機能、アポトーシス関連バイオマーカー、細胞生存率など、複数の指標を同時に検出することを強くお勧めします。カスタマイズされた製品マッチング、詳細な実験プロトコル、または個別技術サポートが必要な場合は、お気軽にお問い合わせください。 Solarbioの専門研究サービスチーム.
FAQ
Q1:酸化ストレスとは具体的に何ですか?
A1:酸化ストレスとは、過剰な活性酸素種(ROS)の生成または抗酸化物質の除去能力の低下によって引き起こされる、細胞内酸化還元恒常性の乱れを指します。この不均衡は、生体分子の損傷、異常なシグナル伝達、炎症反応、アポトーシスによる細胞死、あるいはこれらの病理学的表現型の組み合わせを引き起こします。
Q2:活性酸素種(ROS)は細胞にとって本質的に有害なものですか?
A2:いいえ。低濃度で一時的な活性酸素種(ROS)は、生理学的シグナル伝達物質として重要な役割を果たします。細胞毒性効果を発揮するのは、ROS濃度が持続的に高い場合、または酸化還元感受性細胞小器官(ミトコンドリア、核)内でROSが局所的に過剰になった場合のみです。
Q3:細胞の抗酸化防御において、どの主要なシグナル伝達経路が支配的ですか?
A3:Nrf2-ARE経路は、主要な抗酸化調節軸として機能します。この経路は、SOD、CAT、GSH-Px、グルタチオン依存性合成酵素、および第II相解毒タンパク質の転写を促進し、酸化還元バランスを回復させます。
問題4: NF-κBが酸化ストレス研究において重要な研究対象である理由は?
A4:NF-κBは、ROSの過剰と炎症を結びつける重要な分子的な架け橋です。これは自己増幅型の正のフィードバックループを形成します。ROSがNF-κBを活性化して炎症を誘発し、活性化された免疫細胞がさらにROSを分泌して経路の活性化を維持し、慢性的な組織損傷を悪化させます。
Q5:ROS実験でNACを使用する際に、どのような重要な注意事項を守るべきですか?
A5:研究者は、NACの作用濃度、インキュベーション時間、溶媒組成を事前に最適化し、細胞毒性による干渉を排除するために並行して細胞生存率試験を実施する必要があります。確固たる信頼できる実験結果を得るためには、酸化還元バイオマーカーの多指標同時検出が不可欠です。



