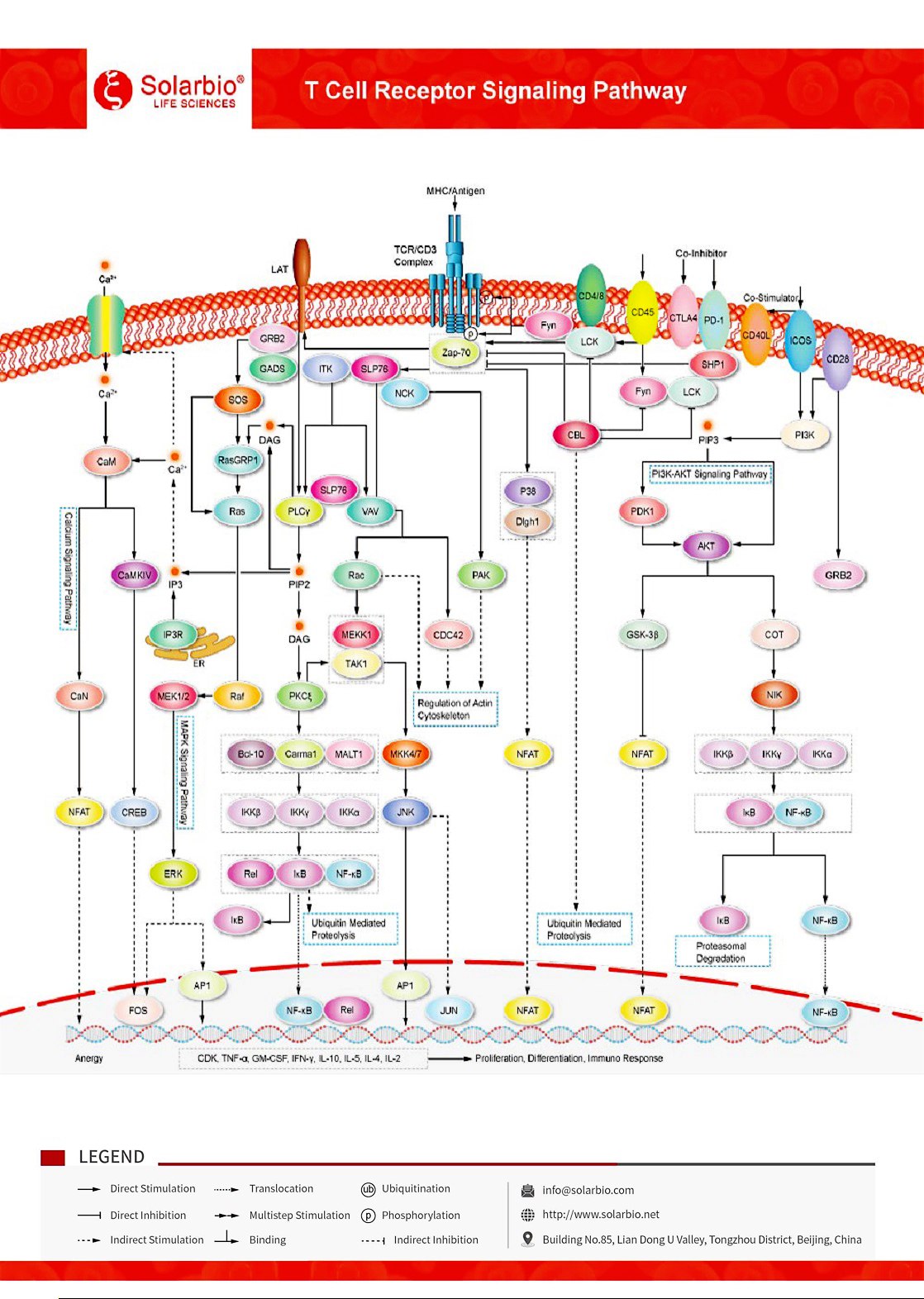





เส้นทางสัญญาณตัวรับเซลล์ T
TCR (ตัวรับเซลล์ T) เป็นเครื่องหมายลักษณะบนพื้นผิวของเซลล์ T ทั้งหมดซึ่งผูกกับ CD3 โดยการเชื่อมโยงที่ไม่มีคุณค่าเพื่อสร้างคอมเพล็กซ์ TCR-CD3 TCR เป็น heterodimer ประกอบด้วย 2 โซ่เปปไทด์ที่แตกต่างกัน α และ β คอมเพล็กซ์ TCR-CD3 ประกอบด้วย β (หรือ γδ) TCR heterodimers ที่หลากหลายทางพันธุกรรมที่ผูกพันกับ CD3 dimers ที่ไม่แตกต่างกัน: CD3ส ส ส ส CD3ส ส ส ส สิ่งที่ไม่แตกต่างกัน: CD3คอคอคอมเซีด TCR เป็นตัวเลือกในการยอมรับชิ้นส่วนเปปไทด์ที่ผูกพันกับโมเลกุลที่ซับซ้อนที่มีความเข้ากันได้กับฮิสโตที่สำคัญบนเซลล์ท การเปิดใช้งานของมันสามารถส่งเสริมการแปลงสัญญาณมากมาย เหตุการณ์แรกของการเปิดใช้งาน TCR คือการฟอสโฟริลของโมติฟการเปิดใช้งานที่ขึ้นอยู่กับตัวรับภูมิคุ้มกันไทโรซิน (ITAMs) บนด้านไซโตพลาสมิกของคอมเพล็กซ์ TCR / CD3 โดยโปรตีนไทโรซินคีนาส (Lck) ของลิมโฟเซ็ต ซึ่งมีโมติฟการเปิดใช้งานที่ขึ้นอยู่กับตัวรับภูมิคุ้มกันไทโร เมื่อฟอสโฟรีเลตโดย Src kinase Lck รูปแบบเหล่านี้เริ่มต้นการส่งสัญญาณเซลล์ T ต่อไป ไทโรซินฟอสฟาเทสของตัวรับ CD45 ควบคุมการฟอสโฟริลและการเปิดใช้งาน Lck และ Src ครอบครัวไทโรซินคินาซ์อื่น ๆ โปรตีนคินาซ (Zap-70) ที่เกี่ยวข้องกับโซ่เซต้า (Zeta-chain) ถูกจ้าง รวมและเปิดใช้งานที่คอมพล็กซ์ TCR / CD3 ซึ่งเริ่มการจ้างและการฟอสโฟริลของอะแดปเตอร์ล่างหรือโปรตีนหลัง เมื่อฟอสโฟรีลโดย Zap-70 SLP-76 ส่งเสริมการรับสมัคร Vav (ปัจจัยการแลกเปลี่ยน guanylate), โปรตีนอะแดปเตอร์ NCK และ GADS และ T-cell kinase (Itk) ที่สามารถอุดมไปได้ การฟอสโฟริเลชั่นของฟอสโฟลิเปซ Cγ1 (PLCγ1) โดย LTK ส่งผลให้เกิดการละลายของฟอสฟาติดิลิโนซิโทล-4, 5-ไดฟอสเฟต (PIP2) เพื่อผลิตส่งข้อความที่สอง diglyceride (DAG) และ inositol triphosphate (IP3) DAG เปิดใช้งานเส้นทาง PKCθ และ MAPK / Erk ซึ่งทั้งสองส่งเสริมกิจกรรมของปัจจัยการเขียน NF-κB IP3 ทริกเกอร์ Ca2+ ปล่อยจาก ER และส่งเสริม Ca นอกเซลล์2+ การเข้าสู่เซลล์ผ่าน Ca2+ ปล่อย Ca ที่เปิดใช้งาน2+(CRAC) ช่อง คา2+ ผูกพัน calmodulin (Ca2+/ CaM) เปิดใช้งาน calcineurin ซึ่งส่งเสริมการถ่ายทอดยีน IL-2 ผ่านปัจจัยการถ่ายทอด NFAT เนื่องจากการศึกษาหลายอย่างได้แสดงให้เห็นว่าระบบการกระตุ้น / การยับยั้งเซลล์ T สามารถใช้เพื่อเพิ่มภูมิคุ้มกันต่อมะเร็ง โมเลกุลหลายอย่างของโปรตีนเหล่านี้กําลังถูกส