




T細胞受容体シグナルパス
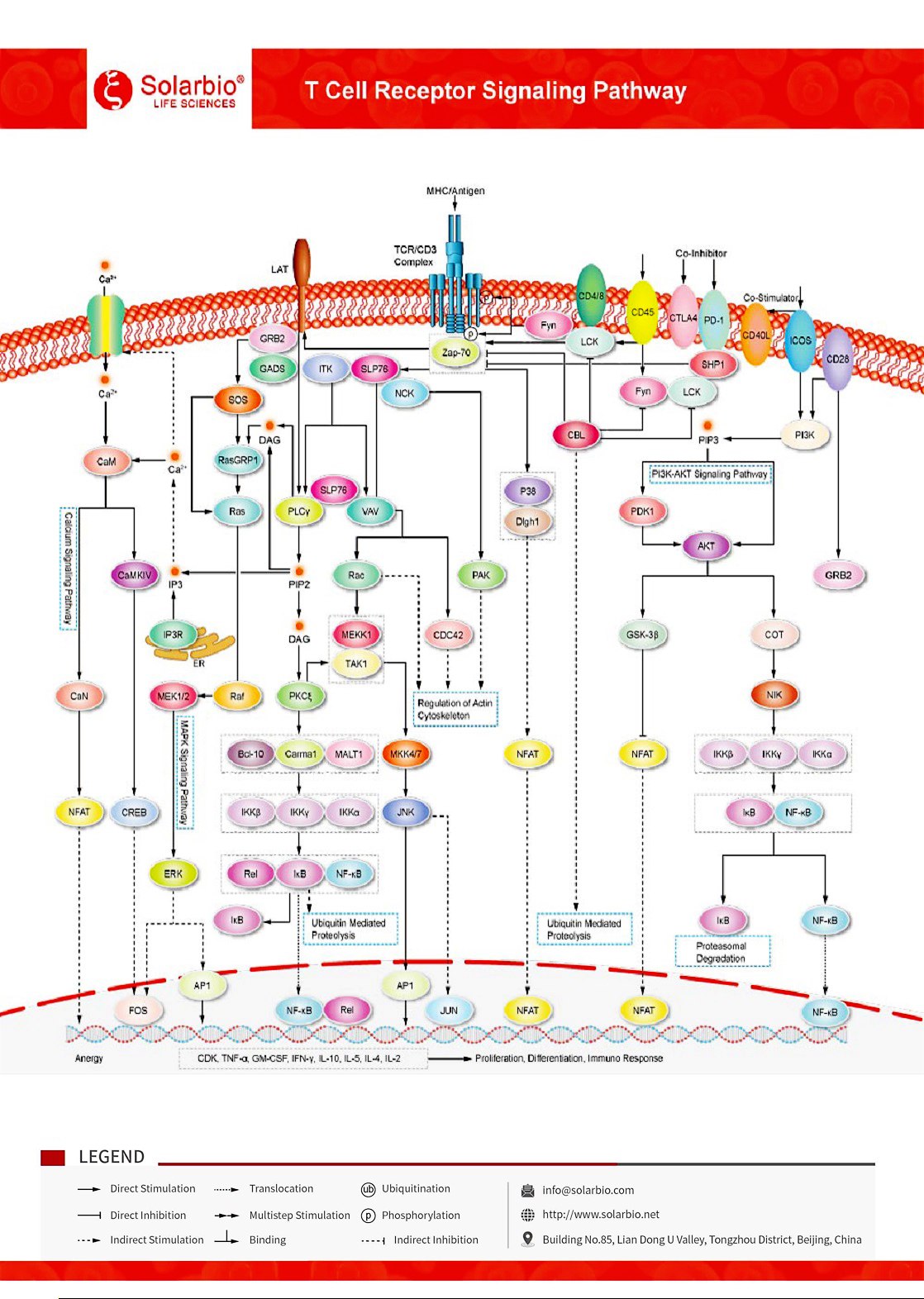
TCR(T細胞受容体)はすべてのT細胞表面の特徴的な標識であり、非共有結合を介してCD 3と結合してTCR−CD 3複合体を形成する。TCRは2つの異なるペプチド鎖αとβからなるイソ二量体である。TCR−CD 3複合体は遺伝的に多様なαβ(またはγδ)TCRイソ二量体から構成され、それらは不変のCD 3二量体と非共有結合結合:CD 3γ、CD 3δ、およびCD 3ゼータゼータである。TCRは、抗原提示細胞上の主要組織適合性複合体分子と結合したペプチド断片を媒介的に識別する。その活性化は多くの信号伝導カスケードを促進することができる。TCR活性化の初期イベントは、リンパ球タンパク質チロシンキナーゼ(Lck)によるTCR/CD 3複合体細胞質側の免疫受容体チロシン依存活性化モチーフ(ITAMs)のリン酸化であり、CD 3ユウロ、γ、δの免疫受容体チロシン依存活性化モチーフ(ITAMs)とゼータ鎖のための免疫受容体チロシンベースの3つの活性化モチーフを含む。SrcキナーゼLckによってリン酸化されると、これらのモチーフは下流のT細胞シグナルカスケードを開始する。CD 45受容体のチロシンホスファターゼは、Lckおよび他のSrcファミリーチロシンキナーゼのリン酸化および活性化を調節する。Zeta鎖関連タンパク質キナーゼ(Zap-70)はTCR/CD 3複合体上で募集、凝集、活性化され、それによって下流アダプタータンパク質または骨格タンパク質の募集とリン酸化を開始する。SLP−76は、Zap−70によってリン酸化されると、Vav(グアニル酸交換因子)、アダプタタンパク質NCKおよびGADS、および誘導T細胞キナーゼ(Itk)の募集を促進する。LTKによるホスホリパーゼCγ1(PLCγ1)のリン酸化によりホスファチジルイノシトール−4,5−二リン酸(PIP 2)が加水分解され、第2メッセンジャージグリセリド(DAG)とイノシトール三リン酸(IP 3)が生成される。DAGはPKCθとMAPK/Erkパスを活性化し、どちらも転写因子NF−κBの活性を促進する。IP 3トリガCa2+ 内質網からの細胞外カルシウムの放出と促進2+ カルシウムを通して細胞に入る2+ かいほうかっせいカルシウム2+(CRAC)チャンネル。カルシウム2+ 結合カルシトニン(Ca2+/CaM)転写因子NFATによりIL−2遺伝子の転写を促進するカルシウム変調神経リン酸酵素を活性化する。多くの研究により、T細胞共刺激/共抑制システムは抗腫瘍免疫を増強するために使用できることが明らかになったため、これらのタンパク質の多くの分子は癌免疫療法の潜在的な標的として研究されている。