




抗体はどんな生体分子ですか。
目次
抗体、免疫グロブリンとも呼ばれ、形質細胞が産生するY形糖蛋白であり、適応免疫の核心である。それらの複雑な4次構造は、可変のグリコシル化モードを加えて、それらを製造および処理条件に特に敏感にする。実験室の科学者にとって、この感受性は持続的な挑戦に転化した:異なる供給業者からの抗体ロットは不一致な蛋白質印影パターン、可変ELISA信号、免疫蛍光中の高度な非特異性背景を生成する可能性がある。厳格な製造検証は極めて重要であるが、ポリクローナル抗体(ポリクローナル免疫反応による固有可変)とモノクローナル抗体または組換え抗体(変異は生産条件と遺伝的ドリフトにより多く発生する)との間の根本的な原因には根本的な違いがある。したがって、この再現性危機を解決するには、より良い品質制御だけでなく、これらの生物学的差異の抗体検証基準を説明する必要がある。
抗体は専門的な糖蛋白質です
構造的には、抗体は2つの同じ重鎖と2つの完全に同じ軽鎖、すなわちκまたはλ同種から構成され、ジスルフィド結合によって特徴的なY形四元構造に連結されている。Yアーム上のFab(フラグメント抗原結合)領域は可変ドメイン(VHとVL)を含み、各ドメインは抗原特異性を付与する相補決定ゾーン(CDR)を3つ有する。Fc(断片結晶性)領域媒介効果器機能、例えば補体活性化及び免疫細胞上のFc受容体と結合し、Asn 297に保存的N−連結グリコシル化部位を担持し、この部位は溶解度、安定性及び抗体依存性細胞毒性(ADCC)を調節する。
この糖タンパク質構造は、なぜ抗体性能が製造条件に非常に敏感であるかを説明する。Fcグリコシル化は宿主細胞グリコシル転移酵素によって触媒される翻訳後修飾であるため、そのパターンはCHO、NS 0、HEK 293細胞などの発現系と培養パラメータに非常に依存している。IgG Fcグリカンは構造的に異質であり、その組成は機能に直接影響する:コアフコースの欠損はFcγRIIIaの結合を増強することによってADCCを増強する、GlcNAcと高ガラクトース(G 2)を2つに分けると補体依存性細胞毒性(CDC)が促進され、Fcグリカン上の末端α2、6−シアル酸はDC−SIGNと結合し、抗炎症作用を発揮する、一方、慢性炎症状態に一般的に存在する無乳糖化(G 0)糖型はCDC低下とFc効果子スペクトル変化を示した。バッチ変異は通常、細胞培養中のグリコシル化の不一致によるものである。したがって、効率的な品質制御は、生産ロットごとの機能的整合性を確保するために、グリカン分析を含むタンパク質濃度測定を超えなければならない。
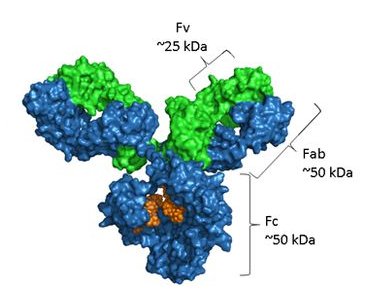
注:青は重鎖、緑は軽鎖、オレンジ部分はグリコシル化を表す。
抗体分類:分子特性を研究ニーズに適合させる
分子レベルで抗体分類を理解することは、賢明な試薬選択を行うことができる:
同種(重鎖変異体)別:
- 免疫グロブリンG:優勢血清抗体(75-80%)、大部分の研究応用の基準、4つのサブクラス(IgG 1-4)、異なる補体活性化能力(IgG 3>IgG 1>IgG 2、IgG 4)、異なるFcγR結合親和力と独特な効果スペクトル、特にIgG 4、sの抗炎症特性と体内Fabアーム交換傾向
- 免疫グロブリンM血清中は五量体を主とし、これは一次免疫反応で産生される第一類抗体である。IgDとともにB細胞受容体(BCR)として発現する場合、モノマー
- 免疫グロブリンA:2つのサブクラス(IgA 1、IgA 2)が存在する、主に粘膜分泌物中のJ鎖媒介重合による二量体である。IgA2;sより短いヒンジ領域は細菌プロテアーゼに対してより大きな抵抗力を持ち、それを腸管微生物群の研究と特に関連させる
- 免疫グロブリンE:単量体、I型超敏感反応と抗ワーム免疫の核心。
- IgD:BCRの一部として、幼稚なB細胞上でIgMと共発現、粘膜安定性および好塩基性粒細胞媒介の免疫調節における分泌機能を認識する人が増えている。
クローン性別:
- モノクローナル抗体(mAbs):ハイブリドーマ技術により単一B細胞クローンから獲得したが、現在も単一B細胞選別、ファージ展示或いは遺伝子組み換え動物プラットフォームにより獲得し、単一エピトープに比類のない特異性を提供した。一貫性のある繰り返し可能な結果を必要とする定量分析(ELISA、フローサイトメトリーなど)に最適です。
- ポリクローナル抗体(pAbs):標的抗原上の複数のエピトープを識別し、強力な信号増幅とより高い変性抗原検出確率を提供し、その中のいくつか(ただしすべてではない)エピトープは完全なままである。免疫組織化学における天然タンパク質の検出、および高親和性シグナル増幅が重要な場合に使用することが好ましい。
宿主種別 &;プロジェクト:
宿主種(ウサギ、マウス、ヤギ、ニワトリ、ラクダ)の選択は、エピトープ識別モード、交差反応特徴、および下流用途との互換性を決定する。組換え抗体およびヒト化抗体は、治療および診断開発のためのキットをさらに拡張する。
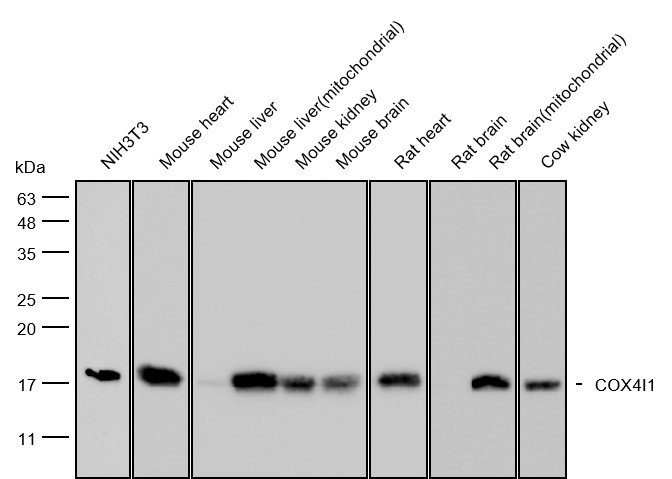
抗体の重要な生物学的機能
コア免疫効果器として、抗体は異なるドメインによって媒介される機能を実行する:
Fab媒介の機能:
- 中和:病原体の宿主細胞への進入を遮断するか、空間障害と受容体競争を通じて毒素活性を中和する。
- ADCC(抗体依存性細胞毒性):FcγRIIIaを介して直接NK細胞が分解抗体包被に関与する標的細胞。
- ADCP(抗体依存性細胞貪食作用):FcγRs媒介マクロファージ貪食の活性化による標的の調整
研究では、抗原結合部位の高い特異性と親和性は、抗体を蛋白質印影、ELISA、IHC、IF、流動細胞法、および共免疫沈殿(co−IP)の不可または欠的な検出ツールにする。治療学では、フコース化によるADCCの増強や突然変異によるサイレント補体活性化などのFc工学戦略が臨床結果を最適化するために積極的に利用されている。
B細胞生物学から一貫性製造へ
体内では、抗体多様性はV(D)J組換えと体細胞高突然変異に由来し、これは適応免疫に必要な過程であるが、研究の再現性要求とは相容れない。この固有の異質性を克服するために、Solarbioは抗体の発見から製造までの制御されたプラットフォームを採用した:
ハイブリドーマ技術:永生化B細胞と骨髄腫が融合し、安定、長期的なモノクローナル抗体分泌を実現する、
組換え哺乳類発現:CHO細胞はグリコシル化スペクトル(グリコシル転移酵素によって調節されたエンジニアリンググリコシルを含む)を制御するために用いられ、HEK 293細胞はヒトグリコシル化を瞬時に生成するために用いられ、
ファージディスプレイ:invitro親和性成熟と抗体断片(scFv、Fabなど)の選択、その後リード線を全長IgGに再フォーマットして組み換え生産を行った。
配列定義のアイデンティティ、結合特異性(ノックアウト/ノックアウトによる細胞系検証)、熱安定性、発現率を含む厳格な先導物選択基準は、極めて重要である。これらの基準を満たしていない抗体は、規模を拡大する際にロット間の可変性を避けられず、どのような生産プラットフォームを用いても、実験の再現性に影響を与える。
研究上の難点:ロット間の可変性
2023年、1000の生命科学実験室を対象とした調査によると、試薬の不一致が免疫測定再現性の失敗の主な原因であることが分かった。表現には弱/欠落したタンパク質印字テープ、ロット交換後のIHC/IFの高背景または信号損失、ELISA標準曲線ドリフトが含まれる。これらの失敗は、検証不足だけでなく、制御されていない製造変数である細胞培養変動、精製不一致、特徴づけられていない糖型異質性に起因する。検証(WB、IHC、IF、フローサイトメトリー、IP)と特異的確認(ノックアウト検証など)を多用しなければ、ある検出に最適化された抗体の別の検出での発現がよくないか、予測不可能なロット間不一致を示すことが多い。
Solarbio抗体検証フレームワーク
北京太陽生物科学技術有限公司、有限会社テクノロジー有限会社は、統合検証パイプラインを通じてこれらの再現性の課題を解決する
- 複数適用相互認証
各抗体は、WB、IHC、IF、FC、IPの5つの標準的な応用によるテストを経て、標的蛋白質とのマッチングによるノックアウト(KO)検証を加えた、生物学的背景。
- 親和性と特異性の定量化
結合強度は表面プラズモン共鳴(SPR)により測定した。相同蛋白質のクロスオーバー反応性スクリーニングにより、脱標的結合を最小限に抑えることができる。
- ロット間一貫性プロトコル
各生産ロットはIPQC、SPC、AQL評価によって追跡される。抗体はアフィニティークロマトグラフィーにより精製し、グリセリンを含む安定剤に供給して凍結融解損傷を防止した。
- 引用文によるパフォーマンスのサポート
Solarbio試薬は、「自然医学」(IF 82.9)や「細胞」(IF 66.85)などの定期刊行物を含む、15,000編を超えるSCIインデックス出版物に引用されている。特異的抗体の応用はリウマチ性関節炎線維芽細胞信号から癌モデルの自食抑制までの研究に現れ、研究者に同業者評議の信頼性証拠を提供した。

研究から反復可能な結果まで
抗体の糖蛋白質としての分類は学術的なものではなく、これらの試薬がどのように製造、検証、貯蔵されなければならないのかを直接知らせ、実験の忠実度を確保する。構造理解とISO認証の品質体系とマルチアプリケーション検証を結合することにより、研究者は免疫測定の信頼性を破壊する固有のロット間の違いを制御することができる。
FAQ
Q 1:抗体はどんな種類の生体分子ですか?
A:糖蛋白(免疫グロブリン)は対をなす重さと軽ポリペプチド鎖から構成され、Fc領域にはN−結合された糖がある。
Q 2:抗体はどのようにして高い特異性を実現しますか?
A:V(D)J組換えと体細胞高突然変異による可変ドメインにおける相補決定領域(CDR)。
Q 3:抗体ロット間の差は何が原因ですか?
A:グリコシル化異質性、凝集/破断及びハイブリドーマクローニングドリフト。厳格な品質制御(糖類分析と機能ロット試験を含む)はこの変異性を制御した。
Q 4:抗体は新規または困難な標的に対してカスタマイズできますか?
A:はい。ハイブリドーマ技術、組換え発現、ファージディスプレイを通じて、研究者は免疫原性の悪い標的(例えば小分子、PTM)、翻訳後修飾または立体特異的エピトープに対する抗体を産生することができる。
Q5:Solarbio抗体は臨床使用に適していますか?
A:いいえ。研究用(RUO)のみです。抗体の生産を研究するには、GMPコンプライアンス、検証されたエンドトキシン限界値、または診断または治療用途に必要な規制ファイルはありません。
|
猫(ねこ) |
製品名 |
文献タイトル |
もし |
|
K107441P |
抗MYH 11ポリクローナル抗体 |
マイクロ流体とマイグレーション学習に基づく複雑な組織高分解能空間分解タンパク質群学 |
45.5 |
|
K008899P |
抗MYH 11ポリクローナル抗体 |
マイクロ流体とマイグレーション学習に基づく複雑な組織高分解能空間分解タンパク質群学 |
45.5 |
|
K106603P |
抗PDIA 3ポリクローナル抗体 |
マイクロ流体とマイグレーション学習に基づく複雑な組織高分解能空間分解タンパク質群学 |
45.5 |
|
0007444p |
抗DCNポリクローナル抗体 |
マイクロ流体とマイグレーション学習に基づく複雑な組織高分解能空間分解タンパク質群学 |
45.5 |
|
K002913P |
抗DCNポリクローナル抗体 |
マイクロ流体とマイグレーション学習に基づく複雑な組織高分解能空間分解タンパク質群学 |
45.5 |
|
K002098P |
抗PDIA 3ポリクローナル抗体 |
マイクロ流体とマイグレーション学習に基づく複雑な組織高分解能空間分解タンパク質群学 |
45.5 |
|
K002095P |
抗MYH 11ポリクローナル抗体 |
マイクロ流体とマイグレーション学習に基づく複雑な組織高分解能空間分解タンパク質群学 |
45.5 |
|
200059 m |
抗β-ミクロ管タンパク質モノクローナル抗体 |
グリボン尿素は代謝調節による見かけの遺伝的修飾によりMDH 2を標的にして老化表現型を緩和する |
40.8 |
|
K109422P |
抗SLP-76ポリクローナル抗体 |
CARに基づく細胞免疫療法のための遺伝的暗号増幅システムによる制御可能で可逆的なスイッチの設計 |
29.5 |
|
K009635P |
抗β-ミクロ管蛋白ポリクローナル抗体 |
移植可能、生分解可能、無線摩擦電気装置、微小管とアクチン動力学の破壊による癌治療 |
29.4 |
|
K009499P |
抗β-ミクロ管蛋白ポリクローナル抗体 |
移植可能、生分解可能、無線摩擦電気装置、微小管とアクチン動力学の破壊による癌治療 |
29.4 |
|
K009362P |
抗β-ミクロ管蛋白ポリクローナル抗体 |
移植可能、生分解可能、無線摩擦電気装置、微小管とアクチン動力学の破壊による癌治療 |
29.4 |
|
200059 m |
抗β-ミクロ管タンパク質モノクローナル抗体 |
移植可能、生分解可能、無線摩擦電気装置、微小管とアクチン動力学の破壊による癌治療 |
29.4 |
|
K106392P |
抗β-ミクロ管蛋白ポリクローナル抗体 |
移植可能、生分解可能、無線摩擦電気装置、微小管とアクチン動力学の破壊による癌治療 |
29.4 |
|
K008723P |
抗RPL 13 Aポリクローナル抗体 |
Cas 13は宿主RNAの内在的な標的化に対してその効用を制限している |
28.1 |
|
K006299P |
抗LY 6 Eポリクローナル抗体 |
Cas 13は宿主RNAの内在的な標的化に対してその効用を制限している |
28.1 |
|
K110275P |
抗−PYCR 3ポリクローナル抗体 |
Cas 13は宿主RNAの内在的な標的化に対してその効用を制限している |
28.1 |
|
K006805P |
抗OASLポリクローナル抗体 |
Cas 13は宿主RNAの内在的な標的化に対してその効用を制限している |
28.1 |
|
K108466P |
抗RBBP 4ポリクローナル抗体 |
腸内GABAエネルギーニューロン由来γ−アミノ酪酸はILC 3の定常状態を維持するためにIgfbp 7の発現を開始する |
27.7 |
|
K200015M |
抗mCherryラベルモノクローナル抗体 |
植物と動物の正義の単鎖RNAウイルスは、その負の義鎖中でウイルス感染に重要な小タンパク質をコードする |
27.5 |
|
200057メートル |
抗GAPDHモノクローナル抗体 |
バイメタル過酸化物ナノ粒子が鉄下垂/銅下垂を一段階共生誘導し、腫瘍免疫治療を強化するためのcGAS STINGパスを活性化する |
27.4 |
|
200058メートル |
抗β-アクチンモノクローナル抗体 |
バイメタル過酸化物ナノ粒子が鉄下垂/銅下垂を一段階共生誘導し、腫瘍免疫治療を強化するためのcGAS STINGパスを活性化する |
27.4 |
|
K114465P |
抗M 6 PRポリクローナル抗体 |
腫瘍の選択的蛋白分解とがん免疫治療の強化のための二機能性リソソーム標的キメラナノプラットフォーム |
27.4 |
|
K009721P |
抗RAGEポリクローナル抗体 |
バイオマーカー活性化された多機能リソソーム標的キメラ媒介細胞外アミロイド繊維の選択的分解 |
23.5 |
|
K001593P |
抗Baxポリクローナル抗体 |
ハイブリッドナノ発電機による閉ループ自己給電低レベル迷走神経刺激システムによる房細動の治療 |
20.577 |
|
K001594P |
抗BCL 2ポリクローナル抗体 |
ハイブリッドナノ発電機による閉ループ自己給電低レベル迷走神経刺激システムによる房細動の治療 |
20.577 |



