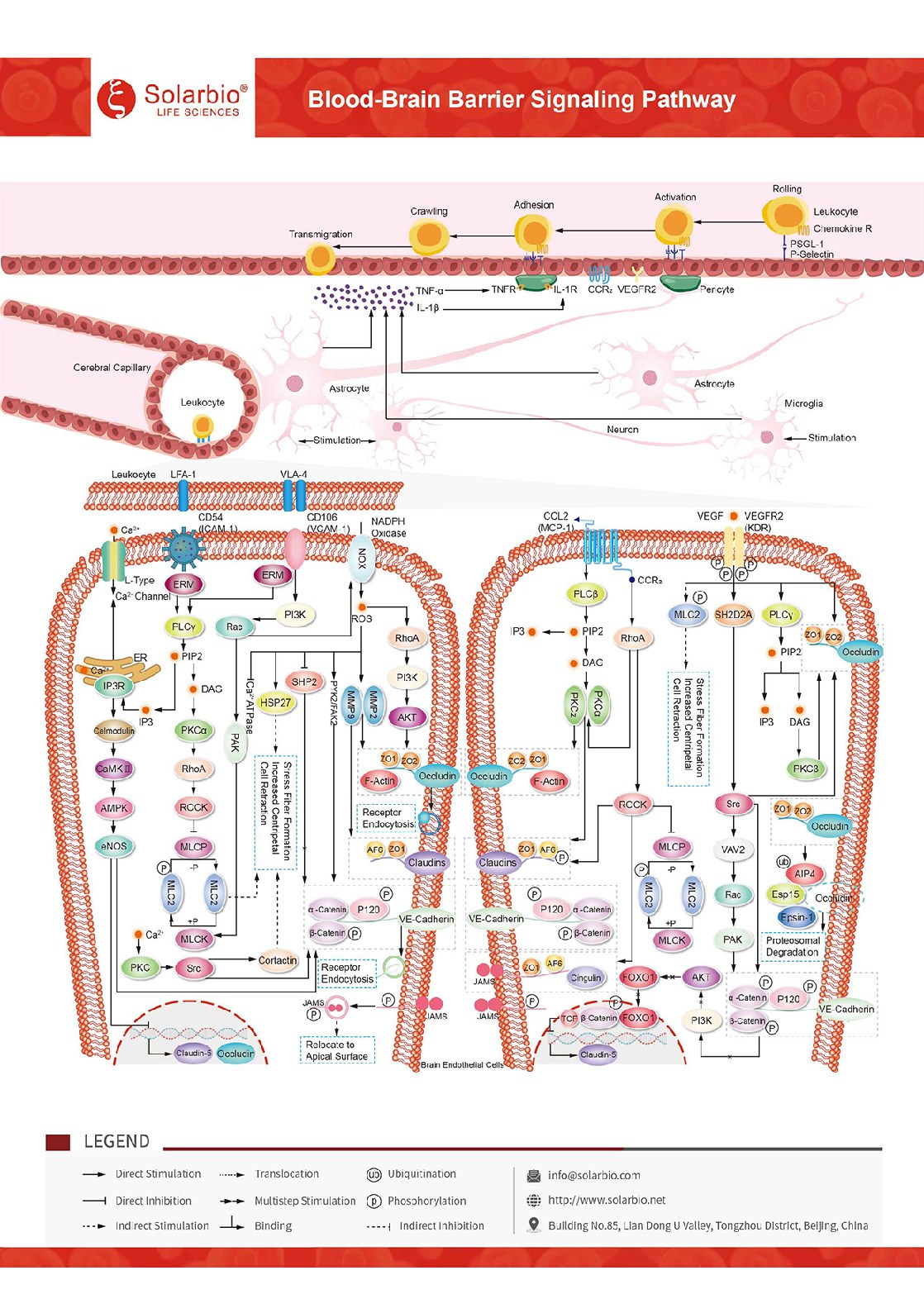





ヘマトクリット信号経路
血液脳障壁は脳微小血管内の強固な障壁である。それは特殊な内皮細胞(EC)からなる物理障壁を含み、先端緊密接続(TJ)、中間接着接続(AJ)と基板外側中部カルシウムチャネルからなる2+−依存性接着、および酵素および排出ポンプタンパク質によって支持される代謝障壁。それらは共に外周循環と中枢神経系(CNS)の間に高度に選択的な拡散障壁を形成する。ケモカインCCL 2/MCP-1は、単核細胞/マクロファージの移動と浸潤を調節する重要なケモカインである。神経炎症状態の中枢神経系では、CCL 2/MCP-1が増加し、脳EC上のGタンパク質カップリング受容体CCR 2と結合する。TJタンパク質のリン酸化を誘導することにより、CCL 2/McP-1は細胞内シグナルカスケードを開始し、細胞縁部からTJタンパク質を分解および/または再分布し、結合複合体の動的再結合とEC収縮をもたらす。JAMタンパク質は内皮細胞の上面に再配向し、インターロイキンとの接着相互作用に関与する。細胞間接着分子−1(ICAM−1/CD 54)は、インターロイキンαLβ2(LFA−1)と結合したIg様接着受容体であり、ICAM−1/CD 5の凝集を誘導し、Srcファミリーキナーゼ及びRho/ROCK経路を含む細胞内信号経路を活性化することができる。血管細胞接着分子−1(VCAM−1/CD 106)はIg様接着受容体であり、インターロイキンα4β1(VLA−4)のリガンドでもある。VCAM−1/CD 106の凝集とTJタンパク質のリン酸化を誘導する細胞内シグナル経路の活性化は、TJタンパク質の細胞縁部からの分解および/または再分布をもたらし、さらに内皮細胞から組織への血液循環による白血球の移行過程を媒介する。また、VCAM-1/CD 106信号伝導はマトリックス金属プロテアーゼ(MMPs)の産生を促進し、マトリックス金属プロテアーゼは結合タンパク質を分解することができる。VEGFは高度に特異的な血管内皮細胞増殖促進因子である。神経炎症状態では、VEGFは中枢神経系で上昇し、脳EC上の受容体と結合し、発生した細胞内シグナルカスケードを起動し、結合複合体の動的組換えとEC収縮を引き起こす。VEGFはOccludinとVE-Cadherinの内呑作用を促進する。あるいは、VEGFは汎素-プロテアーゼ体経路を通じてOccludinの分解を誘導する。VEGFはOccludinとClaudin-5の発現を下方修正することもできる。通常の場合、血液脳障壁の特殊な構造は、大部分の親水性化合物が脳内皮細胞の細胞の傍を通って輸送されるのを阻止し、血液由来細胞の中枢神経系への移動を制限する。神経炎症の過程で、TJとAJなどの結合複合体を再形成して内皮間隙を形成することができ、外周免疫細胞はこの間隙を通じて中枢神経系(CNS)に入る。CCL 2/MCP-1、ICAM-1/CD 54、VCAM-1/CD 106、VEGF信号はいずれもF-actinマイクロフィラメントのストレス繊維への組換えを誘導し、それによって細胞の求心張力を増加させ、細胞収縮を引き起こすことができる。長期的な組織損傷は炎症を引き起こし、血脳関門はその制限機能を失うことになる。反応性小膠細胞、星型膠細胞、周細胞と内皮細胞は大量分子(例えばCXCL 8/IL-8、CCL 2/MCP-1、TNF-α、IL-1β/IL-1 F 2など)を放出し、血管透過性の増加を招いた。それは末梢免疫細胞の中枢神経系への侵入を促進する。報告によると、血脳障壁機能障害とその後の浸潤免疫細胞の増加はアルツハイマー病を含む各種神経系疾患と関係があり、パーキンソン病s病と多発性硬化症。